- Sammensætning C1 kan spalte DNA, der indeholder AP-stedet
- Flere strukturelle dele i C1 er vigtige for katalysen
- DNA-struktur modulerer den C1-katalyserede spaltning
- Spaltning af DNA på AP-steder ved C1 foregår via et mellemprodukt, der involverer den sekundære amin
- C1 øger den termiske stabilitet af DNA, der indeholder en analog af et AP-sted
- C1 har en højere affinitet for DNA indeholdende et AP-sted
- Konklusioner
Sammensætning C1 kan spalte DNA, der indeholder AP-stedet
Den første observation, der antydede tilstedeværelsen af en usædvanlig aktivitet i en delmængde af de undersøgte små molekyler, blev gjort, da C1 (Fig. 1a) blev testet som en potentiel hæmmer af hOGG1. Specifikt resulterede tilføjelse af denne forbindelse til reaktioner af hOGG1 med et fluorescerende mærket duplex oligodeoxynukleotid indeholdende et stedspecifikt 8-oxo-dG addukt (Fig. 1b) i et yderligere produkt, der migrerede hurtigere end β-eliminationsproduktet; hOGG1 alene gav kun β-eliminationsproduktet (Fig. 1c). Der blev ikke observeret nogen nicked-produkter, når 8-oxo-dG-holdigt DNA blev inkuberet med C1 alene.

AP-sted-holdigt DNA blev opnået ved at behandle det tilsvarende dU-holdige DNA med UDG. (a) Struktur af en repræsentativ katalysator til DNA-spaltning. (b) DNA-substrater. (c) Spaltning af 8-oxo-dG-holdigt DNA (250 nM) i tilstedeværelse af hOGG1 (100 nM) og C1 (10 μM). (d) Spaltning af AP-site-holdigt DNA (250 nM) i tilstedeværelse af hOGG1 (50 nM) og C1 (10 μM). (e) Spaltning af AP-site-holdigt DNA (250 nM) i tilstedeværelse af hNEIL1 (50 nM) og C1 (10 μM). (f) Spaltning af AP-site-holdigt DNA (2 μM) ved hjælp af C1. Procentdelen af produkter blev korrigeret for den spontane spaltning. Reaktioner blev udført ved 37 °C i 30 min (b-d) eller 16 h (e).
Der blev opstillet flere mekanistiske muligheder, der kunne forklare dannelsen af det nye produkt. 1) I tilstedeværelse af C1 havde hOGG1 erhvervet evnen til at katalysere en δ-elimineringsreaktion ud over sine glykosylase- og β-eliminerings-AP-lyasefunktioner; 2) C1 kunne konvertere β-eliminationsproduktet til δ-eliminationsproduktet; og 3) AP-steder dannet i glykosylasereaktionen, kunne tjene som et substrat for en C1-katalyseret β,δ-elimineringsreaktion. For at behandle disse muligheder blev duplex-DNA indeholdende et enkelt stedsspecifikt AP-sted genereret ved at behandle det tilsvarende dU-holdige DNA med uracil-DNA-glycosylase (UDG) (Fig. 1b) og reageret med C1 eller hOGG1, enten individuelt eller i kombination. Som med det 8-oxo-dG-holdige oligodeoxynukleotid producerede hOGG1 det forventede β-eliminationsprodukt, mens tilsætning af C1 producerede en blanding af to produkter (fig. 1d). Denne produktblanding blev også dannet, når det AP-holdige oligodeoxynukleotid blev inkuberet med C1 alene. Mens placeringen af det langsommere migrerende produktbånd svarede til β-eliminationsproduktet af hOGG1 (fig. 1d), var det hurtigere migrerende produktbånd sammigreret med det kendte δ-eliminationsprodukt af hNEIL1 (fig. 1e). C1 fremmer således β- og δ-elimineringsreaktionerne på AP-stederne.
Spaltegningsreaktionen ved C1 på AP-sted-holdigt DNA var koncentrationsafhængig (Fig. 1e og Supplerende figur S1), idet lave mikromolære koncentrationer var tilstrækkelige til at observere produkterne. I lighed med de tidligere konstruerede “kunstige nukleaser “26 viste C1 turn-over-katalyse på DNA-substratet: 1 pmol C1 genererede ~1,4 pmol produkter i løbet af 16 timer (fig. 1f). C1’s snithastighed på dette DNA var således ~1,5 × 10-3 min-1 eller højere. Sammenligningen af C1 med de i øjeblikket tilgængelige AP-lyase-reagenser spermine17 og KWKK-peptidet23 viste, at den nyligt identificerede katalysator var mindst 100 gange mere effektiv (Supplerende figur S1).
Flere strukturelle dele i C1 er vigtige for katalysen
Analoger af C1 blev undersøgt for at bestemme bidraget fra specifikke strukturelle træk til strengen spaltningskemien. Vi undersøgte i første omgang evnen af enkle, kommercielt tilgængelige aminer uden indolinon-pyrroldelen C2 (Fig. 2) og CS1-CS3 (Supplerende figur S2) til at skære DNA på et AP-sted. Eventuel dannelse af incisionsprodukter var under detektionsniveauet ved 10 μM. Indolinon-pyrrol-delen er således vigtig for reaktionen. Det er muligt, at indolinon-pyrrol-underenheden binder sig til AP-stedet29 eller minor groove og placerer den sekundære amin til at katalysere strengspaltning via et kovalent mellemprodukt med AP-stedet. Forbindelser C3 (fig. 2) og CS4 (supplerende figur S2), der indeholdt indolinon-pyrrol-enheden, men som manglede den sekundære amin, var også fuldstændig inaktive. Disse data tyder stærkt på, at aminen spiller en rolle som den reaktive funktionelle gruppe. Udskiftning af methoxygruppen på pyrrolet med andre substitutioner, som f.eks. i C4 og C5 (fig. 2), førte til nedsat aktivitet. Reaktionen blev også moduleret af en substituent på aminogruppen, således at en forøgelse af de steriske krav til den sekundære amin mindskede mængden af det observerede produkt (sammenlign C1, C6 og C7 (Fig. 2) og C4, CS5 og CS6 (Supplerende figur S2)). Disse struktur-aktivitetsanalyser viste, at flere af de enkelte dele er nødvendige, men ikke tilstrækkelige til spaltning, og at disse dele skal fungere i samarbejde for at frembringe strengspaltning.

(a) Strukturer af repræsentative forbindelser. (b) DNA-substrat. (c) Test af de repræsentative forbindelsers (10 μM) evne til at skære AP-site-holdigt DNA (250 nM). Reaktioner blev udført ved 37 °C i 30 in.
DNA-struktur modulerer den C1-katalyserede spaltning
Indolinon-pyrrol-delenes betydning for strengen spaltningskemien har antydet, at DNA-substratets struktur kunne påvirke den C1-medierede spaltning. For at undersøge disse forhold blev de indledende reaktionshastigheder målt for et enkeltstrenget DNA (sekvensen som i fig. 1a) og de tilsvarende dobbeltstrengede DNA’er, der indeholdt enten A, C, G eller T over for AP-stedet. Det homopolymeriske oligodeoxynukleotid, 5′-TAMRA-(T)5-AP-(T)11-3′, blev også undersøgt som en entydig enkeltstrenget DNA-struktur. Disse data viste, at selv om strengspaltning kunne finde sted i forbindelse med enkeltstrenget DNA, var dobbeltstrenget DNA de meget foretrukne substrater for C1 (fig. 3). Det blev også konstateret, at arten af basen over for AP-stedet modulerede katalysen, idet hastigheden af AP-stedets hydrolyse var hurtigere over for pyrimidiner end puriner. Disse observationer er i overensstemmelse med forslaget om, at indolinon-pyrrol-underenheden interagerer med DNA for at besætte et tomrum på AP-stedet. Det er bemærkelsesværdigt, at den indledende hastighed, der blev målt for det dobbeltstrengede DNA med et C over for AP-stedet, var meget tæt på den hastighed, der blev observeret for dette substrat under betingelser med begrænset C1-koncentration (fig. 1f).

Reaktioner blev udført ved 37 °C med 250 nM DNA og 5 μM C1. De gennemsnitlige indledende hastigheder med respektive standardafvigelser blev beregnet fra tre uafhængige eksperimenter ved hjælp af KaleidaGraph 4.1-software (Synergy Software). P-værdierne blev beregnet ved hjælp af Students’ t-test.
Spaltning af DNA på AP-steder ved C1 foregår via et mellemprodukt, der involverer den sekundære amin
For at teste for det kovalente mellemprodukt med den ringåbnede, aldehydiske form af deoxyribose, blev et AP-sted-holdigt 32P-mærket oligodeoxynukleotid indeholdende 32P-mærket oligodeoxynukleotid (Fig. 4a) blev inkuberet med C1 i nærværelse af NaB(CN)H3. Selv om dette reduktionsmiddel reagerer langsomt med AP-læsionen, fanger det effektivt imin- eller iminiumion-konjugatet. I en kontrolreaktion med AP-stedet indeholdende oligodeoxynukleotid og NaB(CN)H3 (Fig. 4b, bane 5) udviste en lille brøkdel (~3%) af DNA’et en nedsat mobilitet. Dette produkt med lavt indhold er almindeligt observeret i fældefangstreaktioner23 og repræsenterer sandsynligvis et kompleks med Tris-molekyler, som tidligere vist i reaktioner med malondialdehyd-pyrimidopurinon-DNA-adduktet30. I en positiv kontrolreaktion med lysin-tryptophan-lysin-lysin-lysin-lysin (KWKK)-peptidet23 blev iminintermediæret fanget, hvilket fremgår af forskydningen i DNA-mobiliteten (Fig. 4b, bane 3). I tilstedeværelse af NaB(CN)H3 og C1 dannede størstedelen af DNA’et (~80 %) et kompleks, der optrådte som en art med nedsat mobilitet (Fig. 4b, lane 4). Kompleksdannelsen skyldtes ikke uspecifik binding af C1 til DNA, da der ikke blev observeret nogen forskydning, da det tilsvarende dU-holdige oligodeoxynukleotid blev testet under identiske betingelser (Fig. 4b, vognbane 1). Disse data var i overensstemmelse med hypotesen om, at reagenset var i stand til at danne et kovalent iminiumionintermediat, som derefter placerer en sidegruppe for en protonabstraktion fra sukkerringen.

(a) DNA-substrater. (b) Cyanoborohydridindfangning af et kompleks mellem C1 og AP-site-holdigt DNA. (c) CID-fragmentering af det reducerede iminiumion-intermediat. (d) Fragmentering af det reducerede C1-deoxyribosekonjugat efter enzymatisk fordøjelse.
Den reduktive fældefangst af C1 blev gentaget ved brug af umarkeret AP-sted-holdigt oligodeoxynukleotid (Fig. 4a), og produktet blev analyseret ved massespektrometri (MS). Analysen afslørede en masse, der var i overensstemmelse med det reducerede DNA-C1 kovalente kompleks (m/z 1134,64 for -2). Kollisionsinduceret dissociation (CID) af denne ion resulterede i et komplet sæt af a-B (base)- og w-ioner, hvilket er i overensstemmelse med den reducerede iminium-ion mellem C1 og AP-stedet (Supplerende figur S3). I a-B-ionserien ledsages fragmenteringen af C3′-O-bindingen normalt af det neutrale tab af nukleobasen. Den reducerede binding mellem C1 og AP-stedet forventedes at være mindre labilt, og som følge heraf observerede vi a4-ionen (m/z 1330,1) såvel som a4-B-ionen (m/z 1005,6). Oligodeoxynukleotidet blev også enzymatisk fordøjet og analyseret ved MS (Fig. 4c og Supplerende figur S4). Der blev observeret et fordøjelsesprodukt med en masse, der stemte overens med det reducerede C1-deoxyribosekonjugat (m/z 444,19); fragmentering af denne produktion gav en datterion med m/z 295,09, der var resultatet af det neutrale tab af N-methylamino-2-deoxyribitol (Fig. 4d og supplerende figur S4). Dette produkt var identisk med det produkt, der blev fremstillet fra den reduktive aminationsreaktion af 2-deoxyribose og C1. Disse undersøgelser viser, at C1 kløver AP-sted-holdigt DNA gennem kovalent katalyse, der involverer den sekundære amin.
C1 øger den termiske stabilitet af DNA, der indeholder en analog af et AP-sted
Effekten af C1 blev undersøgt på den termiske stabilitet af DNA, der indeholder en tetrahydrofuran (THF), strukturel analog af et AP-sted, der ikke er i stand til at gennemgå β-elimineringsreaktionen31. Tm for duplex-DNA’et med THF over for T (T:THF) steg med 6,3 °C ved tilsætning af 1 ækvivalent C1, mens Tm for kontrol-DNA’et med et T:A-par var mindre påvirket og kun steg med 1 °C (fig. 5). Mens smeltekurven for T:THF med C1 indikerede termisk stabilisering ved hjælp af C1, udvidede tilstedeværelsen af C1 også smeltekurven, hvilket indikerer en mindre samarbejdsvillig smelteovergang. Samlet set tyder disse observationer på, at C1 specifikt binder sig til DNA på AP-stedet, hvilket giver en lokaliseret stabilisering af DNA-helixen. Dette er i overensstemmelse med den foreslåede model for interaktioner mellem C1 og DNA, hvor indolinon-pyrrol-delen optager den plads, der er tilgængelig på AP-stedet.
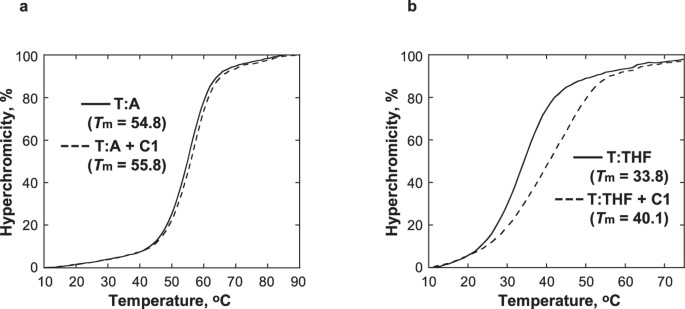
Smeltetemperaturkurverne for T:A (a) og T:THF (b) blev opnået ved brug af 5 μM DNA i 10 mM natriumphosphatbuffer, pH 7.0, 100 mM NaCl og 1 mM EDTA i fravær (gennemgående linje) og tilstedeværelse (stiplet linje) af 5 μM C1.
C1 har en højere affinitet for DNA indeholdende et AP-sted
For at undersøge C1’s bindingsmåde og affinitet til AP-sted-holdigt DNA blev der udført cirkulær dichroisme (CD)-analyser ved hjælp af både THF-holdige og kontrol T:A duplex oligodeoxynukleotider (Fig. 6). Den inducerede CD (ICD) af C1 ved interaktion med T:THF-duplex blev observeret som et stærkt excitonsignal, en bisignatform med positive og negative bånd i forhold til det frie C1’s absorptionsmaksimum. Dette er generelt tegn på dannelse af dimeriske komplekser eller komplekser af højere orden, enten i en rillebindende eller en ekstern stabelbindende tilstand32. Titreringen af C1 med stigende T:THF-koncentrationer (fig. 6a og supplerende figur S5) afslørede både uspecifikke og specifikke interaktioner. Ved titreringens start er C1 i overskud, og uspecifikke interaktioner er begunstiget. Ved tilsætning af T:THF steg ICD’en ved ~488 nm. Ved ca. 2 μM DNA-koncentration skiftede ICD-båndet til lavere energi (~495 nm) og faldt derefter indtil ækvivalenspunktet og plateauede i overskud af DNA, hvilket indikerer en specifik interaktion. Titrering af C1 med kontrol-T:A-DNA (fig. 6a og supplerende figur S5) viste ikke det positive ICD-bånd omkring 495 nm, hvilket indikerer fraværet af specifikke interaktioner. Den fremadrettede titrering af T:THF viste også tegn på uspecifikke og specifikke interaktioner (fig. 6b). Ved lave koncentrationer af C1 med DNA i overskud var ICD-båndet centreret ved ~495 nm (specifikt); efterhånden som koncentrationen af C1 steg i forhold til DNA, steg ICD-intensiteten og skiftede til højere energi (~488 nm), hvilket indikerer uspecifik binding som forventet ved høje ligandkoncentrationer. Det observerede hypsokromiske skift kan indikere en ændring i eller en mere defineret konformation af C1, når den er specifikt bundet til AP-stedet. Titrering af kontrol-DNA’et uden AP-sted viste også en ICD og tegn på uspecifik binding (fig. 6c). Det er værd at bemærke, at båndet ved 450-520 nm kun optrådte ved et 3-dobbelt overskud af C1 i forhold til DNA, hvilket gør dette bølgelængdeområde til det bedste valg til at konstruere bindingskurverne. Bindingsisothermer blev konstrueret og analyseret ved hjælp af en simpel bimolekylær bindingsmodel som tidligere offentliggjort32. De ikke-lineære ligninger, der fremkom af 500 nm isotermerne (Fig. 6d), gav en dissociationskonstant (KD) på 64 μM for kontrol-T:A-DNA’et (uspecifik binding) og 29 μM for T:THF-DNA’et (specifik binding); KD for C1 med THF-holdigt duplex blev beregnet til 22 μM efter subtraktion for at korrigere for det uspecifikke bindingsbidrag og mulige ligand-ligand-interaktioner (tabel 1). KD-værdierne beregnet ved 495 nm lå kun lidt uden for fejlene i forhold til værdierne beregnet ved 500 nm (tabel 1).

(a) Titreringskurver af ICD-intensiteten ved 500 nm ved anvendelse af en konstant C1-koncentration (20 μM) og stigende koncentrationer af DNA (omvendt titrering). (b) CD-spektre for T:THF og (c) T:A ved anvendelse af en konstant DNA-koncentration (10 μM) og stigende koncentrationer af C1 (fremadrettet titrering). (d) Titreringskurver for ICD-intensiteten ved 500 nm over for logaritmen af C1-koncentrationen for T:THF og T:A (afledt af dataene i panelerne b og c). (e) Titreringskurver for ICD-intensitet ved 500 nm over for logaritmen af C6-koncentrationen for T:THF og T:A (afledt af dataene i supplerende figur S6). (d) Titreringskurver af ICD-intensiteten ved 500 nm over for logaritmen af C6-koncentrationen for T:AP og T:A (afledt af dataene i supplerende figur S6).
Der er endvidere blevet overvejet den mulighed, at affiniteten af C1 kunne være anderledes for DNA, der indeholder et naturligt AP-sted i stedet for THF. For at besvare dette spørgsmål testede vi den strukturelt beslægtede men inaktive forbindelse C6 (fig. 2) i CD-analyser med DNA, der enten indeholder THF eller et AP-sted, der blev skabt fra dU ved UDG-behandling. De CD-spektre, der blev observeret for C6 i fremadrettede titreringseksperimenter med T:THF- og T:A-DNA’er (Supplerende figur S6), lignede generelt dem, der blev observeret for C1 (fig. 6b,c). Bindingsisothermerne blev konstrueret (fig. 6e) og analyseret som ovenfor. KD for C6 med THF-holdigt duplex, korrigeret for de uspecifikke interaktioner, blev beregnet til 28 μM (tabel 1), hvilket kun er ~25 % højere end den tilsvarende KD, der blev beregnet for C1. C6 synes således at være en passende model til undersøgelse af C1’s interaktioner med DNA. Derefter blev der udført forward-titrering af C6 ved hjælp af DNA, der indeholdt det UDG-afledte AP-site (Supplerende figur S6). Analyser af bindingsisotermer viste, at C6’s affinitet til dette DNA var stort set identisk med THF-holdigt DNA (fig. 6f og tabel 1). I betragtning af, at deoxyribose på et naturligt AP-sted overvejende findes i den ring-lukkede, THF-lignende form31 , var sidstnævnte resultat ikke overraskende. Man kan forvente, at affiniteten af C1 til AP-site-holdigt DNA før dannelsen af et kovalent mellemprodukt ville være sammenlignelig med den affinitet, der er målt for THF-holdigt DNA. Den stærkere binding af C1 til DNA, der indeholder et AP-sted, sammenlignet med kontrol-DNA’et var således i overensstemmelse med dataene om termisk stabilitet og understøtter tilsammen forslaget om, at indolinon-pyrrol-underenheden besætter et tomt rum på AP-stedet i DNA’et. Det er sandsynligt, at C1’s affinitet for AP-stedet bidrager til dets lyaseaktivitet og giver en klar fordel i forhold til C2, CS1, CS2 og CS3, som mangler indolinon-pyrrol-delen (Fig. 2 og Supplerende figur S2).
Konklusioner
De AP DNA-strengsplitningskatalysatorer, der er beskrevet i denne undersøgelse, repræsenterer en ny klasse af forbindelser, der kan udnyttes til at spalte AP-steder under fysiologiske forhold. Baseret på denne kernestruktur kan molekyler designes til at forbedre selektiviteten af AP-stedskæring over for forskellige baser og potentielt for forskellige sekvenskontekster. På grund af de seneste fremskridt i udviklingen af screeningsteknikker med højt gennemløb kan disse forbindelser hurtigt udvikles til at blive effektive reagenser til at kløve AP-steder, der er skabt ved depurination eller BER-vejen i celler og organismer. De optimerede versioner af C1 kan også have terapeutiske anvendelser, især i kombination med mange almindelige kræftmidler, der enten skader DNA, såsom alkylerende midler33 , eller er rettet mod DNA-reparation, såsom PARP34,35 eller AP-endonuklease36 -inhibitorer. Det er værd at nævne, at de 3′-ender, der er skabt via β- eller β,δ-elimination, ikke kan udnyttes af DNA-polymeraser uden forudgående reparation37. De biologiske konsekvenser af omdannelsen af AP-steder til sådanne DNA-strengbrud forventes at være komplekse og varierer afhængigt af den cellulære evne til at reparere eller tolerere forskellige typer DNA-skader. De mulige resultater kan omfatte en øget terapeutisk effektivitet af behandlinger mod kræft (mere effektiv celledrab) og en mindsket lægemiddelinduceret mutagenese. Spaltning af DNA på AP-steder kan være særlig gavnlig for behandlingen af kræftformer med defekter i mekanismerne til reparation af DNA-strengebrud, f.eks. kræftformer med BRCA-mangel34,35.