- Il composto C1 può scindere il DNA contenente il sito AP
- Diverse società strutturali in C1 sono importanti per la catalisi
- La struttura del DNA modula la scissione catalizzata da C1
- La scissione del DNA nei siti AP da parte di C1 procede attraverso un intermedio che coinvolge l’ammina secondaria
- C1 aumenta la stabilità termica del DNA contenente un analogo di un sito AP
- C1 ha una maggiore affinità per il DNA contenente un sito AP
- Conclusioni
Il composto C1 può scindere il DNA contenente il sito AP
L’osservazione iniziale che ha suggerito la presenza di un’attività insolita in un sottoinsieme delle piccole molecole in studio è stata fatta quando C1 (Fig. 1a) è stato provato come potenziale inibitore di hOGG1. In particolare, l’aggiunta di questo composto alle reazioni di hOGG1 con un oligodeossinucleotide marcato con fluorescenza contenente un addotto site-specific 8-oxo-dG (Fig. 1b) ha portato ad un prodotto aggiuntivo che è migrato più velocemente del prodotto di β-eliminazione; hOGG1 da solo ha prodotto solo il prodotto di β-eliminazione (Fig. 1c). Nessun prodotto intaccato è stato osservato quando il DNA contenente 8-oxo-dG è stato incubato con C1 da solo.

Il DNA contenente il sito AP è stato ottenuto trattando il corrispondente DNA contenente dU con UDG. (a) Struttura di un catalizzatore rappresentativo per la scissione del DNA. (b) Substrati di DNA. (c) Scissione del DNA contenente 8-oxo-dG (250 nM) in presenza di hOGG1 (100 nM) e C1 (10 μM). (d) Scissione del DNA contenente il sito AP (250 nM) in presenza di hOGG1 (50 nM) e C1 (10 μM). (e) Scissione del DNA contenente il sito AP (250 nM) in presenza di hNEIL1 (50 nM) e C1 (10 μM). (f) Scissione del DNA contenente il sito AP (2 μM) da C1. La percentuale di prodotti è stata corretta per la scissione spontanea. Le reazioni sono state effettuate a 37 °C per 30 min (b-d) o 16 h (e).
Sono state ipotizzate diverse possibilità meccaniche che potrebbero spiegare la formazione del nuovo prodotto. 1) In presenza di C1, hOGG1 ha acquisito la capacità di catalizzare una reazione di δ-eliminazione oltre alle sue funzioni di glicosilasi e di β-eliminazione AP liasi; 2) C1 potrebbe convertire il prodotto di β-eliminazione nel prodotto di δ-eliminazione; e 3) i siti AP formati nella reazione di glicosilasi, potrebbero servire come substrato per una reazione di β,δ-eliminazione catalizzata da C1. Per affrontare queste possibilità, il DNA duplex contenente un singolo sito specifico AP è stato generato trattando il corrispondente DNA contenente dU con uracile DNA glicosilasi (UDG) (Fig. 1b) e reagito con C1 o hOGG1, individualmente o in combinazione. Come con l’oligodeossinucleotide contenente 8-oxo-dG, hOGG1 ha prodotto il prodotto atteso di β-eliminazione, mentre l’aggiunta di C1 ha prodotto una miscela di due prodotti (Fig. 1d). Questa miscela di prodotti si è formata anche quando l’oligodeossinucleotide contenente AP è stato incubato con C1 da solo. Mentre la posizione della banda del prodotto che migra più lentamente corrispondeva al prodotto di β-eliminazione di hOGG1 (Fig. 1d), la banda del prodotto che migra più velocemente si è co-migrata con il prodotto di δ-eliminazione noto di hNEIL1 (Fig. 1e). Così, C1 promuove le reazioni di β- e δ-eliminazione nei siti AP.
La reazione di scissione da C1 sul DNA contenente il sito AP era dipendente dalla concentrazione (Fig. 1e e Figura supplementare S1), con basse concentrazioni micromolari sufficienti per osservare i prodotti. Simile alle “nucleasi artificiali” precedentemente progettate 26, C1 ha mostrato una catalisi turn-over sul substrato di DNA: 1 pmol C1 generato ~ 1,4 pmol prodotti in 16 h (Fig. 1f). Così, il tasso di incisione da C1 su questo DNA era ~ 1,5 × 10-3 min-1 o superiore. Il confronto di C1 con i reagenti di liasi AP attualmente disponibili spermina17 e il peptide KWKK23 ha dimostrato che il catalizzatore appena identificato era almeno 100 volte più efficiente (Figura supplementare S1).
Diverse società strutturali in C1 sono importanti per la catalisi
Analoghi di C1 sono stati testati per determinare il contributo di specifiche caratteristiche strutturali alla chimica di scissione del filamento. Abbiamo inizialmente esaminato la capacità di semplici ammine disponibili in commercio prive della frazione C2 dell’indolinone-pirrolo (Fig. 2) e CS1-CS3 (Figura supplementare S2) per incidere il DNA in un sito AP. Formazione di prodotti di incisione, se del caso, era sotto il livello di rilevamento a 10 μM. Così, l’indolinone-pirrolo moiety è importante per la reazione. E ‘possibile che l’indolinone-pirrolo subunità si lega al sito AP 29 o solco minore e posiziona l’ammina secondaria per catalizzare la scissione del filo attraverso un intermedio covalente con il sito AP. I composti C3 (Fig. 2) e CS4 (Figura supplementare S2) che contenevano la parte indolinone-pirrolo ma senza l’ammina secondaria erano anche completamente inattivi. Questi dati hanno fortemente suggerito il ruolo dell’ammina come gruppo funzionale reattivo. La sostituzione del gruppo metossi sul pirrolo con altre sostituzioni, come in C4 e C5 (Fig. 2), ha portato a una diminuzione dell’attività. La reazione è stata modulata anche da un sostituente sul gruppo amminico, in modo tale che aumentando le richieste steriche dell’ammina secondaria è diminuita la quantità di prodotto osservato (confrontare C1, C6, e C7 (Fig. 2), e C4, CS5, e CS6 (Figura supplementare S2)). Queste analisi struttura-attività hanno dimostrato che diverse delle singole società sono necessarie ma non sufficienti per la scissione, e che queste società devono agire in modo cooperativo per produrre la scissione del filamento.

(a) Strutture di composti rappresentativi. (b) Substrato di DNA. (c) Saggio per la capacità dei composti rappresentativi (10 μM) di incidere il DNA contenente il sito AP (250 nM). Le reazioni sono state effettuate a 37 °C per 30 in.
La struttura del DNA modula la scissione catalizzata da C1
L’importanza della frazione indolinone-pirrolo per la chimica di scissione del filamento ha suggerito che la struttura del substrato di DNA potrebbe influenzare la scissione mediata da C1. Per affrontare queste relazioni, i tassi iniziali di reazione sono stati misurati per un DNA a singolo filamento (la sequenza come in Fig. 1a) e i corrispondenti DNA a doppio filamento che contenevano A, C, G, o T di fronte al sito AP. L’oligodeossinucleotide omopolimerico, 5′-TAMRA-(T)5-AP-(T)11-3′, è stato anche esaminato come una struttura univoca di DNA a singolo filamento. Questi dati hanno dimostrato che, sebbene la scissione dei filamenti possa avvenire nel contesto del DNA a singolo filamento, i DNA a doppio filamento erano i substrati preferiti per C1 (Fig. 3). Si è anche scoperto che la natura della base opposta al sito AP modulava la catalisi, con il tasso di idrolisi del sito AP più veloce di fronte alle pirimidine che alle purine. Queste osservazioni sono coerenti con la proposta che la subunità indolinone-pirrolo interagisce con il DNA per occupare uno spazio vuoto nel sito AP. In particolare, il tasso iniziale misurato per il DNA a doppio filamento con una C di fronte al sito AP, era molto vicino al tasso osservato per questo substrato in condizioni di concentrazione limitante C1 (Fig. 1f).

Le reazioni sono state effettuate a 37 °C usando 250 nM di DNA e 5 μM di C1. I tassi medi iniziali con le rispettive deviazioni standard sono stati calcolati da tre esperimenti indipendenti utilizzando il software KaleidaGraph 4.1 (Synergy Software). I valori P sono stati calcolati usando il test t degli studenti.
La scissione del DNA nei siti AP da parte di C1 procede attraverso un intermedio che coinvolge l’ammina secondaria
Per testare l’intermedio covalente con l’anello aperto, forma aldeidica del desossiribosio, un oligodeossinucleotide marcato con 32P nel sito AP (Fig. 4a) è stato incubato con C1 in presenza di NaB(CN)H3. Mentre questo riduttore reagisce lentamente con la lesione AP, intrappola efficacemente lo ione imina o iminio coniugato. In una reazione di controllo con il sito AP contenente oligodeossinucleotide e NaB (CN) H3 (Fig. 4b, corsia 5), una piccola frazione (~ 3%) del DNA ha manifestato una mobilità diminuita. Questo prodotto a bassa abbondanza è comunemente osservato nelle reazioni di intrappolamento 23 e probabilmente rappresenta un complesso con molecole Tris, come è stato precedentemente dimostrato nelle reazioni con l’addotto malondialdeide pirimidopurinone DNA 30. In una reazione di controllo positivo utilizzando la lisina-triptofano-lisina-lisina (KWKK) peptide 23, l’intermedio imina è stato intrappolato, come evidenziato dallo spostamento della mobilità del DNA (Fig. 4b, corsia 3). In presenza di NaB(CN)H3 e C1, la maggior parte del DNA (~80%) ha formato un complesso che è apparso come una specie con mobilità diminuita (Fig. 4b, corsia 4). La formazione del complesso non era dovuta al legame non specifico di C1 al DNA, poiché non è stato osservato alcuno spostamento quando il corrispondente oligodeossinucleotide contenente dU è stato testato in condizioni identiche (Fig. 4b, corsia 1). Questi dati erano coerenti con l’ipotesi che il reagente fosse in grado di formare un intermedio covalente di ione iminio, che poi posiziona un gruppo laterale per un’astrazione di protoni dall’anello di zucchero.

(a) substrati di DNA. (b) intrappolamento di cianoboroidride di un complesso tra C1 e il DNA contenente il sito AP. (c) Frammentazione CID dell’intermedio ridotto dello ione iminio. (d) frammentazione del coniugato ridotto C1-deossiribosio dopo la digestione enzimatica.
L’intrappolamento riduttivo di C1 è stato ripetuto utilizzando senza etichetta AP sito contenente oligodeossinucleotide (Fig. 4a), e il prodotto è stato analizzato dalla spettrometria di massa (MS). L’analisi ha rivelato una massa coerente con il complesso covalente ridotto DNA-C1 (m/z 1134.64 per -2). Dissociazione indotta da collisione (CID) di questo ione ha portato in una serie completa di a-B (Base) e w ioni, coerente con lo ione iminium ridotto intermedio tra C1 e il sito AP (Figura supplementare S3). Nella serie di ioni a-B, la frammentazione del legame C3′-O è normalmente accompagnata dalla perdita neutra della nucleobase. Il legame ridotto tra C1 e il sito AP ci si aspettava che fosse meno labile e, di conseguenza, abbiamo osservato lo ione a4 (m/z 1330.1) così come il a4-B (m/z 1005.6). L’oligodeossinucleotide è stato anche digerito enzimaticamente e analizzato da MS (Fig. 4c e Figura supplementare S4). Un prodotto di digestione è stato osservato con una massa coerente con il coniugato ridotto C1-deossiribosio (m / z 444.19), la frammentazione di questo ione prodotto ha dato uno ione figlia con m / z 295.09, che è risultato dalla perdita neutro di N-metil amino-2-deossiribitolo (Fig. 4d e Figura supplementare S4). Questo prodotto era identico a quello preparato dalla reazione di amminazione riduttiva di 2-deossiribosio e C1. Questi studi dimostrano che C1 taglia il DNA contenente un sito AP attraverso la catalisi covalente che coinvolge l’ammina secondaria.
C1 aumenta la stabilità termica del DNA contenente un analogo di un sito AP
L’effetto di C1 è stato studiato sulla stabilità termica del DNA contenente un tetraidrofurano (THF), analogo strutturale di un sito AP che è incapace di subire la reazione di β-eliminazione31. La Tm del DNA duplex contenente THF opposto T (T:THF) è aumentato di 6,3 ° C con l’aggiunta di 1 equivalente di C1, mentre la Tm del DNA di controllo contenente una coppia T:A era meno influenzato e solo aumentato di 1 ° C (Fig. 5). Mentre la curva di fusione di T:THF con C1 indicato stabilizzazione termica da C1, la presenza di C1 anche ampliato la curva di fusione, indicando una transizione meno cooperativa fusione. Collettivamente, queste osservazioni suggeriscono che C1 si lega specificamente al DNA nel sito AP fornendo una stabilizzazione localizzata all’elica del DNA. Questo è coerente con il modello proposto di interazioni tra C1 e il DNA in cui la moiety indolinone-pirrolo occupa lo spazio disponibile nel sito AP.
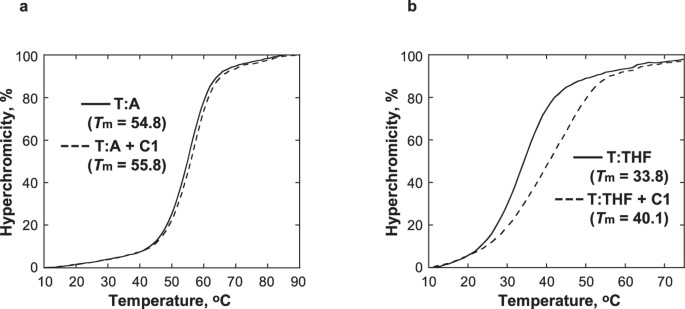
Le curve della temperatura di fusione di T:A (a) e T:THF (b) sono state ottenute usando 5 μM di DNA in 10 mM di tampone fosfato di sodio, pH 7.0, 100 mM NaCl, e 1 mM EDTA in assenza (linea continua) e presenza (linea tratteggiata) di 5 μM C1.
C1 ha una maggiore affinità per il DNA contenente un sito AP
Al fine di esaminare la modalità di legame e l’affinità di C1 al DNA contenente un sito AP, sono state condotte analisi di dicroismo circolare (CD) utilizzando sia oligodeossinucleotidi contenenti THF che duplex T:A di controllo (Fig. 6). Il CD indotto (ICD) di C1 dopo l’interazione con il duplex T:THF è stato osservato come un forte segnale di eccitoni, una forma bisignata con bande positive e negative relative al massimo di assorbimento del C1 libero. Questo è generalmente indicativo della formazione di complessi dimerici o di ordine superiore, sia in una modalità di groove-binding o un esterno stacking-binding32. La titolazione di C1 con crescenti concentrazioni di T:THF (Fig. 6a e Figura supplementare S5) ha rivelato entrambe le interazioni aspecifiche e specifiche. All’inizio della titolazione, C1 è in eccesso e interazioni aspecifiche sono favoriti. Con l’aggiunta di T:THF, l’ICD a ~ 488 nm è aumentato. A circa 2 μM concentrazione di DNA, la banda ICD spostato a bassa energia (~ 495 nm) e poi diminuito fino al punto di equivalenza e plateaued in eccesso di DNA che indica una interazione specifica. La titolazione di C1 con il controllo T:A DNA (Fig. 6a e Figura supplementare S5) non ha mostrato l’ICD positivo intorno a 495 nm indicando l’assenza di interazioni specifiche. La titolazione in avanti di T:THF ha anche mostrato prove di interazioni aspecifiche e specifiche (Fig. 6b). A basse concentrazioni di C1 con il DNA in eccesso, la banda ICD è stata centrata a ~ 495 nm (specifico); come la concentrazione di C1 aumentato rispetto al DNA, l’intensità ICD aumentato e spostato a più alta energia (~ 488 nm), indicativo di legame non specifico come previsto ad alte concentrazioni di ligando. Lo spostamento ipocromico osservato può indicare un cambiamento o una conformazione più definita di C1 quando è specificamente legato al sito AP. Titolazione del DNA di controllo senza un sito AP anche mostrato un ICD e la prova di legame non specifico (Fig. 6c). Vale la pena notare che la banda a 450-520 nm è apparso solo ad un eccesso di 3 volte di C1 rispetto al DNA, rendendo questo intervallo di lunghezza d’onda la scelta migliore per costruire le curve di legame. Isoterme di legame sono stati costruiti e analizzati utilizzando un semplice modello di legame bimolecolare come pubblicato in precedenza 32. Le equazioni non lineari che sono risultati dalle isoterme 500 nm (Fig. 6d) ha dato la costante di dissociazione (KD) di 64 micron per il controllo T: A DNA (legame aspecifico) e 29 micron per il T: THF DNA (legame specifico), il KD di C1 con THF contenente duplex è stato calcolato per essere 22 micron dopo la sottrazione per correggere il contributo non specifico legame e possibili ligando-ligando interazioni (Tabella 1). I valori KD calcolati a 495 nm erano solo leggermente al di fuori degli errori di quelli calcolati a 500 nm (Tabella 1).

(a) Curve di titolazione dell’intensità ICD a 500 nm utilizzando una concentrazione costante di C1 (20 μM) e concentrazioni crescenti di DNA (titolazione inversa). (b) spettri CD per T:THF e (c) T:A usando una concentrazione costante di DNA (10 μM) e concentrazioni crescenti di C1 (titolazione in avanti). (d) Curve di titolazione di intensità ICD a 500 nm contro il logaritmo della concentrazione di C1 per T:THF e T:A (derivato dai dati in pannelli b e c). (e) Curve di titolazione di intensità ICD a 500 nm contro il logaritmo della concentrazione di C6 per T:THF e T:A (derivato dai dati in Figura supplementare S6). (d) Curve di titolazione dell’intensità ICD a 500 nm rispetto al logaritmo della concentrazione di C6 per T:AP e T:A (derivate dai dati della Figura S6 supplementare).
Inoltre, è stata considerata la possibilità che l’affinità di C1 possa essere diversa per il DNA contenente un sito AP naturale invece del THF. Per rispondere a questa domanda, abbiamo testato il composto strutturalmente correlato ma inattivo C6 (Fig. 2) in analisi CD con DNA contenente THF o un sito AP creato da dU mediante trattamento UDG. Gli spettri CD osservati per C6 in esperimenti di titolazione in avanti utilizzando il T:THF e T:A DNAs (Figura supplementare S6) era generalmente simile a quello osservato per C1 (Fig. 6b,c). Le isoterme di legame sono stati costruiti (Fig. 6e) e analizzati come sopra. Il KD di C6 con THF contenente duplex, corretto per le interazioni non specifiche è stato calcolato per essere 28 μM (Tabella 1), che è solo ~ 25% superiore al KD corrispondente calcolato per C1. Così, C6 sembra essere un modello appropriato per studiare le interazioni di C1 con il DNA. Titolazione in avanti di C6 è stata poi eseguita utilizzando il DNA che conteneva il UDG-derivato sito AP (Figura supplementare S6). Le analisi delle isoterme di legame hanno rivelato che l’affinità di C6 a questo DNA era essenzialmente identica al DNA contenente THF (Fig. 6f e Tabella 1). Considerando che deossiribosio in un sito naturale AP esiste prevalentemente in anello chiuso, THF-come forma 31, quest’ultimo risultato non era sorprendente. Si può anticipare che prima della formazione di un intermedio covalente, l’affinità di C1 a AP sito contenente DNA sarebbe paragonabile a quella misurata per THF contenente DNA. Quindi, il legame più forte di C1 al DNA contenente un sito AP rispetto al DNA di controllo è stato coerente con i dati di stabilità termica e insieme supporta la proposta che la subunità indolinone-pirrolo occupa uno spazio vuoto nel sito AP del DNA. È probabile che l’affinità di C1 per il sito AP contribuisca alla sua attività di liasi e fornisca un chiaro vantaggio rispetto a C2, CS1, CS2 e CS3, a cui manca la parte indolinone-pirrolo (Fig. 2 e Figura supplementare S2).
Conclusioni
I catalizzatori di scissione del filamento di DNA AP descritti in questo studio rappresentano una nuova classe di composti che possono essere utilizzati per tagliare i siti AP in condizioni fisiologiche. Sulla base di questa struttura centrale, le molecole possono essere progettate per migliorare la selettività dell’incisione del sito AP di fronte a diverse basi e potenzialmente per diversi contesti di sequenza. Grazie ai recenti progressi nello sviluppo di tecniche di screening ad alta produttività, questi composti potrebbero essere rapidamente avanzati per essere reagenti efficienti per scindere i siti AP che sono creati attraverso la depurinazione o il percorso BER in cellule e organismi. Le versioni ottimizzate di C1 potrebbero anche avere applicazioni terapeutiche, in particolare in combinazione con molti farmaci antitumorali comuni che danneggiano il DNA, come gli agenti alchilanti33, o mirano alla riparazione del DNA, come gli inibitori della PARP34,35 o dell’endonucleasi AP36 . Vale la pena ricordare che le estremità 3′ create tramite β- o β,δ-eliminazione non possono essere utilizzati da DNA polimerasi senza riparazione preliminare37. Le conseguenze biologiche della conversione dei siti AP in tali rotture del filamento di DNA dovrebbero essere complesse e variare a seconda della capacità cellulare di riparare o tollerare diversi tipi di danni al DNA. I possibili risultati possono includere una maggiore efficacia terapeutica dei trattamenti anti-cancro (uccisione più efficiente delle cellule) e una diminuzione della mutagenesi indotta dai farmaci. La scissione del DNA nei siti AP può essere particolarmente vantaggiosa per il trattamento dei tumori che hanno difetti nei meccanismi di riparazione delle rotture del filamento di DNA, come i tumori con carenza di BRCA34,35.
.