- Compound C1 can cleave DNA containing AP site
- Verschillende structurele moieties in C1 zijn belangrijk voor de katalyse
- DNA-structuur moduleert de C1-gematalyseerde splitsing
- Cleavage van DNA op AP-plaatsen door C1 verloopt via een tussenproduct waarbij het secundaire amine betrokken is
- C1 verhoogt de thermische stabiliteit van DNA dat een analoog van een AP-locatie bevat
- C1 heeft een hogere affiniteit voor DNA dat een AP-locatie bevat
- Conclusies
Compound C1 can cleave DNA containing AP site
De eerste waarneming die de aanwezigheid van een ongebruikelijke activiteit in een subset van de bestudeerde kleine moleculen suggereerde, werd gedaan toen C1 (Fig. 1a) werd getest als een potentiële remmer van hOGG1. Toevoeging van deze verbinding aan reacties van hOGG1 met een fluorescent gelabeld duplex oligodeoxynucleotide dat een plaatsspecifiek 8-oxo-dG adduct bevat (Fig. 1b) resulteerde in een extra product dat sneller migreerde dan het β-eliminatieproduct; hOGG1 alleen leverde alleen het β-eliminatieproduct (Fig. 1c). Er werden geen ingeknaagde producten waargenomen wanneer 8-oxo-dG-bevattend DNA met C1 alleen werd geïncubeerd.

AP-locatie-bevattend DNA werd verkregen door het overeenkomstige dU-bevattend DNA met UDG te behandelen. (a) Structuur van een representatieve katalysator voor DNA-splitsing. (b) DNA-substraten. (c) Splijting van 8-oxo-dG-bevattend DNA (250 nM) in aanwezigheid van hOGG1 (100 nM) en C1 (10 μM). (d) Splijting van AP site-bevattend DNA (250 nM) in aanwezigheid van hOGG1 (50 nM) en C1 (10 μM). (e) Splitsing van AP site-bevattend DNA (250 nM) in aanwezigheid van hNEIL1 (50 nM) en C1 (10 μM). (f) Splitsing van AP site-bevattend DNA (2 μM) door C1. Het percentage producten is gecorrigeerd voor de spontane splijting. De reacties werden uitgevoerd bij 37 °C gedurende 30 min (b-d) of 16 uur (e).
Er werden verschillende mechanistische mogelijkheden verondersteld die de vorming van het nieuwe product zouden kunnen verklaren. 1) In aanwezigheid van C1 had hOGG1 het vermogen verworven om naast zijn glycosylase- en β-eliminatie-AP lyase-functies ook een δ-eliminatiereactie te katalyseren; 2) C1 kon het β-eliminatieproduct omzetten in het δ-eliminatieproduct; en 3) AP-locaties die in de glycosylase-reactie waren gevormd, konden dienen als substraat voor een C1-gekatalyseerde β,δ-eliminatiereactie. Om deze mogelijkheden te onderzoeken werd duplex-DNA dat een enkele plaats-specifieke AP-plaats bevatte gegenereerd door het overeenkomstige dU-bevattende DNA te behandelen met uracil-DNA-glycosylase (UDG) (fig. 1b) en gereageerd met C1 of hOGG1, afzonderlijk of in combinatie. Net als bij het 8-oxo-dG-bevattende oligodeoxynucleotide produceerde hOGG1 het verwachte β-eliminatieproduct, terwijl de toevoeging van C1 een mengsel van twee producten opleverde (fig. 1d). Dit productmengsel werd ook gevormd wanneer het AP-bevattende oligodeoxynucleotide met C1 alleen werd geïncubeerd. Terwijl de positie van de langzamer migrerende productband overeenkwam met het β-eliminatieproduct van hOGG1 (fig. 1d), co-migreerde de sneller migrerende productband met het bekende δ-eliminatieproduct van hNEIL1 (fig. 1e). Aldus bevordert C1 de β- en δ-eliminatiereacties op AP-locaties.
De splitsingsreactie door C1 op AP-locatie-bevattend DNA was concentratieafhankelijk (fig. 1e en supplementair figuur S1), waarbij lage micromolaire concentraties voldoende waren om de producten waar te nemen. Vergelijkbaar met de eerder ontworpen “kunstmatige nucleasen “26, C1 toonde turn-over katalyse op het DNA-substraat: 1 pmol C1 genereerde ~1.4 pmol producten in 16 uur (Fig. 1f). De incisiesnelheid van C1 op dit DNA was dus ~1,5 × 10-3 min-1 of hoger. De vergelijking van C1 met de momenteel beschikbare AP lyase reagentia spermine17 en het KWKK peptide23 toonde aan dat de nieuw-geïdentificeerde katalysator ten minste 100-voudig efficiënter was (Supplementary Figure S1).
Verschillende structurele moieties in C1 zijn belangrijk voor de katalyse
Analogen van C1 werden onderzocht om de bijdrage van specifieke structurele kenmerken aan de chemische splitsing van de streng te bepalen. In eerste instantie onderzochten wij het vermogen van eenvoudige, in de handel verkrijgbare amines zonder het indolinon-pyrrol-gedeelte C2 (fig. 2) en CS1-CS3 (aanvullend figuur S2) om DNA op een AP-locatie te insnijden. Vorming van incisie producten, indien aanwezig, was onder het niveau van detectie bij 10 pM. Het indolinon-pyrrool gedeelte is dus belangrijk voor de reactie. Het is mogelijk dat de indolinon-pyrrool subunit bindt aan de AP site29 of kleine groef en de secundaire amine positioneert om streng splitsing te katalyseren via een covalent intermediair met de AP site. Verbindingen C3 (Fig. 2) en CS4 (Supplementary Figure S2) die het indolinon-pyrrool gedeelte bevatten maar het secundaire amine misten waren ook volledig inactief. Deze gegevens suggereerden sterk de rol van het amine als de reactieve functionele groep. De vervanging van de methoxygroep op de pyrrol door andere substituties, zoals in C4 en C5 (Fig. 2), leidde tot verminderde activiteit. De reactie werd ook gemoduleerd door een substituent op de aminogroep, zodanig dat het verhogen van de sterische eisen van het secundaire amine de hoeveelheid van het waargenomen product verminderde (vergelijk C1, C6, en C7 (Fig. 2), en C4, CS5, en CS6 (Supplementary Figure S2)). Deze structuur-activiteitsanalyses toonden aan dat verschillende van de afzonderlijke mootjes noodzakelijk maar niet voldoende zijn voor splitsing, en dat deze mootjes moeten samenwerken om splitsing van de streng te bewerkstelligen.

(a) Structuren van representatieve verbindingen. (b) DNA-substraat. (c) Bepaling van het vermogen van representatieve verbindingen (10 μM) om AP-locatie-bevattend DNA (250 nM) in te snijden. De reacties werden uitgevoerd bij 37 °C gedurende 30 in.
DNA-structuur moduleert de C1-gematalyseerde splitsing
Het belang van het indolinon-pyrrol-gedeelte voor de chemische splitsing van de streng heeft gesuggereerd dat de structuur van het DNA-substraat van invloed zou kunnen zijn op de C1-gemedieerde splitsing. Om deze verbanden te onderzoeken werden de aanvankelijke reactiesnelheden gemeten voor een enkelstrengs DNA (de sequentie als in fig. 1a) en de corresponderende dubbelstrengs DNA’s die ofwel A, C, G, of T tegenover de AP plaats bevatten. Het homopolymere oligodeoxynucleotide, 5′-TAMRA-(T)5-AP-(T)11-3′, werd ook onderzocht als een ondubbelzinnige enkelstrengs DNA-structuur. Deze gegevens toonden aan dat, hoewel strengafsplitsing in de context van enkelstrengs DNA kon plaatsvinden, de dubbelstrengs DNA’s de sterk geprefereerde substraten voor C1 waren (fig. 3). Ook werd vastgesteld dat de aard van de base tegenover de AP-locatie de katalyse moduleerde, waarbij de snelheid van hydrolyse van de AP-locatie tegenover pyrimidines sneller was dan tegenover purines. Deze waarnemingen zijn consistent met het voorstel dat de indolinon-pyrrol-subeenheid interageert met DNA om een lege ruimte op de AP-locatie in te nemen. Opmerkelijk is dat de beginsnelheid gemeten voor het dubbelstrengs DNA met een C tegenover de AP-plaats, zeer dicht bij de snelheid lag die voor dit substraat werd waargenomen onder omstandigheden van beperkende C1-concentratie (Fig. 1f).

Reacties werden uitgevoerd bij 37 °C met 250 nM DNA en 5 μM C1. De gemiddelde beginsnelheden met de respectieve standaardafwijkingen werden berekend op basis van drie onafhankelijke experimenten met behulp van KaleidaGraph 4.1-software (Synergy Software). De P-waarden werden berekend met behulp van Students’ t-test.
Cleavage van DNA op AP-plaatsen door C1 verloopt via een tussenproduct waarbij het secundaire amine betrokken is
Om te testen of er een covalent tussenproduct is met de ringgeopende, aldehydische vorm van de deoxyribose, werd een AP-plaats-bevattend 32P gelabeld oligodeoxynucleotide (Fig. 4a) werd geïncubeerd met C1 in aanwezigheid van NaB(CN)H3. Hoewel dit reductans langzaam reageert met de AP-laesie, vangt het het imine- of iminiumion-conjugaat efficiënt op. In een controlereactie met het AP-locatie-bevattende oligodeoxynucleotide en NaB(CN)H3 (fig. 4b, laan 5) vertoonde een kleine fractie (~3%) van het DNA een verminderde mobiliteit. Dit product met een lage abundantie wordt vaak waargenomen in de vangstreacties23 en vertegenwoordigt waarschijnlijk een complex met Tris-moleculen, zoals eerder is aangetoond in reacties met het malondialdehyde pyrimidopurinon DNA-adduct30. In een positieve controlereactie met het lysine-tryptofaan-lysine-lysine (KWKK)-peptide23 werd het imine-tussenproduct gevangen, zoals blijkt uit de verschuiving in de DNA-mobiliteit (Fig. 4b, laan 3). In aanwezigheid van NaB(CN)H3 en C1 vormde het merendeel van het DNA (~80%) een complex dat verscheen als een soort met verminderde mobiliteit (Fig. 4b, laan 4). De complexvorming was niet het gevolg van niet-specifieke binding van C1 aan DNA, aangezien er geen verschuiving werd waargenomen toen het overeenkomstige dU-bevattende oligodeoxynucleotide onder identieke omstandigheden werd getest (fig. 4b, laan 1). Deze gegevens waren consistent met de hypothese dat het reagens in staat was een covalent iminiumiontussenproduct te vormen, dat vervolgens een zijgroep positioneert voor een protonabstractie van de suikerring.

(a) DNA-substraten. (b) Cyanoborohydride-vangst van een complex tussen C1 en AP-site-bevattend DNA. (c) CID fragmentatie van het gereduceerde iminium ion intermediair. (d) Fragmentatie van het gereduceerde C1-deoxyriboseconjugaat na enzymatische digestie.
De reductieve vangst van C1 werd herhaald met gebruikmaking van ongelabeld AP-locatie-bevattend oligodeoxynucleotide (fig. 4a), en het product werd geanalyseerd met massaspectrometrie (MS). De analyse toonde een massa die consistent is met het gereduceerde DNA-C1 covalente complex (m/z 1134.64 voor -2). Botsing geïnduceerde dissociatie (CID) van dit ion resulteerde in een complete reeks van a-B (Base) en w ionen, consistent met het gereduceerde iminium ion intermediair tussen C1 en de AP site (Supplementary Figuur S3). In de a-B-ionenreeks gaat fragmentatie van de C3′-O-binding normaal gesproken gepaard met het neutrale verlies van de nucleobase. Verwacht werd dat de verminderde binding tussen C1 en de AP-locatie minder labiel zou zijn en als gevolg daarvan namen we zowel het a4-ion (m/z 1330,1) als het a4-B-ion (m/z 1005,6) waar. Het oligodeoxynucleotide werd ook enzymatisch ontsloten en geanalyseerd met MS (Fig. 4c en Supplementary Figure S4). Er werd een digestieproduct waargenomen met een massa die consistent was met het gereduceerde C1-deoxyriboseconjugaat (m/z 444,19); fragmentatie van dit production gaf een dochterion met m/z 295,09, dat het resultaat was van het neutrale verlies van N-methyl amino-2-deoxyribitol (fig. 4d en aanvullende figuur S4). Dit product was identiek aan het product dat werd bereid uit de reductieve amineringreactie van 2-deoxyribose en C1. Deze studies tonen aan dat C1 AP-locatie-bevattend DNA splitst door covalente katalyse waarbij het secundaire amine betrokken is.
C1 verhoogt de thermische stabiliteit van DNA dat een analoog van een AP-locatie bevat
Het effect van C1 werd bestudeerd op de thermische stabiliteit van DNA dat een tetrahydrofuraan (THF), structureel analoog van een AP-locatie bevat die niet in staat is de β-eliminatiereactie te ondergaan31. De Tm van het duplex-DNA met THF tegenover T (T:THF) nam met 6,3 °C toe na toevoeging van 1 equivalent C1, terwijl de Tm van het controle-DNA met een T:A-paar minder werd beïnvloed en slechts met 1 °C toenam (Fig. 5). Terwijl de smeltcurve van T:THF met C1 duidde op thermische stabilisatie door C1, verbreedde de aanwezigheid van C1 ook de smeltcurve, wat duidt op een minder coöperatieve smeltovergang. Tezamen suggereren deze waarnemingen dat C1 zich specifiek bindt aan DNA op de AP-locatie, waardoor de DNA-helix plaatselijk wordt gestabiliseerd. Dit is consistent met het voorgestelde model van interacties tussen C1 en DNA waarbij het indolinon-pyrrool-gedeelte de beschikbare ruimte op de AP-plaats inneemt.
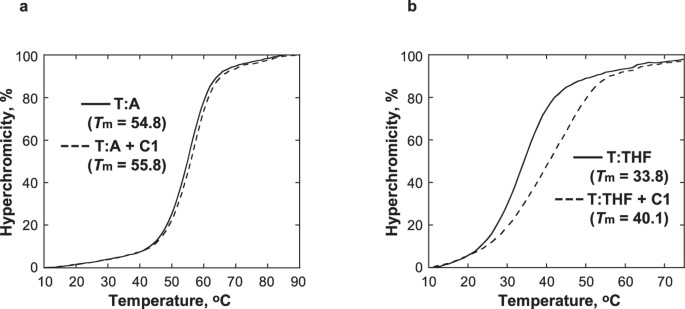
De smelttemperatuurcurven van T:A (a) en T:THF (b) werden verkregen met 5 μM DNA in 10 mM natriumfosfaatbuffer, pH 7.0, 100 mM NaCl, en 1 mM EDTA in afwezigheid (volle lijn) en aanwezigheid (stippellijn) van 5 μM C1.
C1 heeft een hogere affiniteit voor DNA dat een AP-locatie bevat
Om de bindingswijze en affiniteit van C1 voor AP-locatie-bevattend DNA te onderzoeken, werden analyses van circulair dichroïsme (CD) uitgevoerd met zowel THF-bevattende als controle T:A duplex oligodeoxynucleotiden (Fig. 6). De geïnduceerde CD (ICD) van C1 bij interactie met T:THF duplex werd waargenomen als een sterk excitonsignaal, een bisignaatvorm met positieve en negatieve banden ten opzichte van het absorptiemaximum van het vrije C1. Dit wijst in het algemeen op de vorming van dimere complexen of complexen van hogere orde, hetzij in een groefbindende, hetzij in een externe stapelingsbindende modus32. De titratie van C1 met toenemende T:THF-concentraties (fig. 6a en aanvullende figuur S5) bracht zowel niet-specifieke als specifieke interacties aan het licht. Aan het begin van de titratie is C1 in overmaat en zijn niet-specifieke interacties in het voordeel. Bij toevoeging van T:THF nam de ICD bij ~488 nm toe. Bij ongeveer 2 μM DNA-concentratie verschoof de ICD-band naar een lagere energie (~495 nm) en daalde vervolgens tot het equivalentiepunt en plateau in overmaat aan DNA, wat duidt op een specifieke interactie. Titratie van C1 met het T:A controle-DNA (fig. 6a en aanvullende figuur S5) vertoonde niet de positieve ICD rond 495 nm, wat duidt op de afwezigheid van specifieke interacties. De voorwaartse titratie van T:THF vertoonde ook aanwijzingen voor niet-specifieke en specifieke interacties (fig. 6b). Bij lage concentraties van C1 met DNA in overmaat was de ICD-band gecentreerd bij ~495 nm (specifiek); naarmate de concentratie van C1 ten opzichte van DNA toenam, nam de ICD-intensiteit toe en verschoof deze naar een hogere energie (~488 nm), wat duidt op niet-specifieke binding zoals verwacht bij hoge ligandconcentraties. De waargenomen hypsochromische verschuiving kan duiden op een verandering in of een meer gedefinieerde conformatie van C1 wanneer deze specifiek aan de AP-locatie is gebonden. Titratie van het controle-DNA zonder AP-locatie toonde ook een ICD en aanwijzingen voor niet-specifieke binding (fig. 6c). Opgemerkt dient te worden dat de band bij 450-520 nm alleen verscheen bij een 3-voudige overmaat van C1 ten opzichte van DNA, waardoor dit golflengtegebied de beste keuze was om de bindingskrommen te construeren. Bindingsisothermen werden geconstrueerd en geanalyseerd met behulp van een eenvoudig bimoleculair bindingsmodel zoals eerder gepubliceerd32. De niet-lineaire vergelijkingen die resulteerden uit de 500 nm isothermen (fig. 6d) gaven een dissociatieconstante (KD) van 64 μM voor het controle T:A DNA (niet-specifieke binding) en 29 μM voor het T:THF DNA (specifieke binding); de KD van C1 met THF-bevattend duplex werd berekend op 22 μM na aftrek om te corrigeren voor de niet-specifieke bindingsbijdrage en mogelijke ligand-ligand interacties (tabel 1). De KD-waarden berekend bij 495 nm vielen slechts iets buiten de fouten van die berekend bij 500 nm (tabel 1).

(a) Titratiecurven van de ICD-intensiteit bij 500 nm met gebruikmaking van een constante C1-concentratie (20 μM) en toenemende concentraties DNA (omgekeerde titratie). (b) CD-spectra voor T:THF en c) T:A bij een constante DNA-concentratie (10 μM) en stijgende concentraties C1 (voorwaartse titratie). (d) Titratiekrommen van de ICD-intensiteit bij 500 nm versus de logaritme van de C1-concentratie voor T:THF en T:A (afgeleid van de gegevens in de panelen b en c). (e) Titratiekrommen van de ICD-intensiteit bij 500 nm versus de logaritme van de C6-concentratie voor T:THF en T:A (afgeleid van de gegevens in supplementaire figuur S6). (d) Titratiecurves van de ICD-intensiteit bij 500 nm versus de logaritme van de C6-concentratie voor T:AP en T:A (afgeleid van de gegevens in aanvullende figuur S6).
Verder is de mogelijkheid overwogen dat de affiniteit van C1 anders zou kunnen zijn voor DNA dat een natuurlijke AP-locatie bevat in plaats van THF. Om deze vraag te beantwoorden, hebben wij de structureel verwante maar inactieve verbinding C6 (fig. 2) in CD-analyses getest met DNA dat THF bevat of een AP-locatie die door middel van UDG-behandeling uit dU is gemaakt. De CD-spectra die voor C6 werden waargenomen in voorwaartse titratie-experimenten met de T:THF en T:A DNA’s (aanvullend Figuur S6) waren over het algemeen vergelijkbaar met die voor C1 (Fig. 6b,c). De bindingsisothermen werden geconstrueerd (fig. 6e) en geanalyseerd zoals hierboven. De KD van C6 met THF-bevattend duplex, gecorrigeerd voor de niet-specifieke interacties, werd berekend op 28 μM (tabel 1), hetgeen slechts ~25% hoger is dan de corresponderende KD berekend voor C1. C6 lijkt dus een geschikt model te zijn voor het bestuderen van interacties van C1 met DNA. Voorwaartse titratie van C6 werd vervolgens uitgevoerd met DNA dat de van UDG afgeleide AP-locatie bevatte (aanvullend figuur S6). Analyses van bindingsisothermen toonden aan dat de affiniteit van C6 voor dit DNA in wezen identiek was aan THF-bevattend DNA (fig. 6f en tabel 1). Gezien het feit dat desoxyribose op een natuurlijke AP-locatie voornamelijk bestaat in de ring-gesloten, THF-achtige vorm31 , was dit laatste resultaat niet verrassend. Verwacht kan worden dat voorafgaand aan de vorming van een covalent tussenproduct de affiniteit van C1 voor AP-locatie-bevattend DNA vergelijkbaar zou zijn met de affiniteit die is gemeten voor THF-bevattend DNA. De sterkere binding van C1 aan DNA dat een AP-locatie bevat in vergelijking met het controle-DNA was dus consistent met de thermische stabiliteitsgegevens en ondersteunt samen het voorstel dat de indolinon-pyrrol-subeenheid een lege ruimte op de AP-locatie van DNA bezet. Het is waarschijnlijk dat de affiniteit van C1 voor de AP plaats bijdraagt aan zijn lyase-activiteit en een duidelijk voordeel biedt ten opzichte van C2, CS1, CS2, en CS3, die het indolinon-pyrrool gedeelte missen (Fig. 2 en Supplementary Figure S2).
Conclusies
De in deze studie beschreven AP DNA strengsplijtingskatalysatoren vertegenwoordigen een nieuwe klasse verbindingen die kunnen worden gebruikt om AP plaatsen onder fysiologische omstandigheden te splitsen. Gebaseerd op deze kernstructuur, kunnen moleculen worden ontworpen om de selectiviteit van AP-locatie-insnijding tegenover verschillende basen en mogelijk voor verschillende sequentiecontexten te verbeteren. Dankzij recente vooruitgang in de ontwikkeling van high-throughput screeningtechnieken zouden deze verbindingen snel kunnen worden ontwikkeld tot efficiënte reagentia om AP-locaties te klieven die ontstaan door depurinatie of de BER-route in cellen en organismen. De geoptimaliseerde versies van C1 kunnen ook therapeutische toepassingen hebben, met name in combinatie met veel gangbare antikankermedicijnen die ofwel DNA beschadigen, zoals alkylerende middelen33 , of gericht zijn op DNA-reparatie, zoals PARP34,35 of AP endonuclease36 remmers. Het is vermeldenswaard dat de 3′-uiteinden die via β- of β,δ-eliminatie tot stand komen, niet door DNA-polymerasen kunnen worden gebruikt zonder voorafgaande reparatie37. De biologische gevolgen van de omzetting van AP-plaatsen in dergelijke DNA-strengbreuken zullen naar verwachting complex zijn en variëren naar gelang van de cellulaire capaciteit om verschillende soorten DNA-schade te repareren of te tolereren. Mogelijke resultaten zijn een verhoogde therapeutische werkzaamheid van anti-kankerbehandelingen (efficiëntere celdoding) en verminderde door geneesmiddelen veroorzaakte mutagenese. De splitsing van DNA op AP-plaatsen kan bijzonder gunstig zijn voor de behandeling van kankers die defecten vertonen in mechanismen voor het herstel van DNA-strengbreuken, zoals BRCA-deficiënte kankers34,35.