- C1 comprimido pode clivar DNA contendo AP sítio
- Moieties estruturais transversais em C1 são importantes para a catálise
- A estrutura do DNA modula a clivagem C1-catalítica
- Cleavage of DNA at AP sites by C1 proceeds via an intermediate involving the secondary amine
- C1 aumenta a estabilidade térmica do DNA contendo um análogo de um sítio AP
- C1 tem maior afinidade para DNA contendo um sítio AP
- Conclusões
C1 comprimido pode clivar DNA contendo AP sítio
A observação inicial que sugeriu a presença de uma atividade incomum em um subconjunto das pequenas moléculas em estudo foi feita quando C1 (Fig. 1a) foi testado como um inibidor potencial do hOGGG1. Especificamente, a adição deste composto às reações do hOGG1 com um oligodeoxinucleotídeo duplex rotulado fluorescentemente contendo um aduto 8-oxo-dG específico do local (Fig. 1b) resultou em um produto adicional que migrou mais rapidamente do que o produto de eliminação β; só o hOGG1 produziu apenas o produto de eliminação β (Fig. 1c). Nenhum produto danificado foi observado quando o DNA contendo 8-oxo-dG foi incubado apenas com C1.

DNA contendo AP foi obtido através do tratamento do DNA contendo dU correspondente com UDG. (a) Estrutura de um catalisador representativo para a cisão do ADN. (b) Substratos de ADN. (c) Cisão de DNA contendo 8-oxo-dG (250 nM) na presença de hOGG1 (100 nM) e C1 (10 μM). (d) Cisão de ADN contendo AP (250 nM) na presença de hOGG1 (50 nM) e C1 (10 μM). (e) Cisão de AP site contendo DNA (250 nM) na presença de hNEIL1 (50 nM) e C1 (10 μM). (f) Cisão de AP site contendo ADN (2 μM) por C1. A porcentagem de produtos foi corrigida para a clivagem espontânea. Reações foram realizadas a 37 °C durante 30 min (b-d) ou 16 h (e).
Possibilidades mecanicistas transversais que poderiam ser responsáveis pela formação do novo produto. 1) Na presença do C1, o hOGG1 tinha adquirido a capacidade de catalisar uma reação de eliminação de δ, além das funções da glicosilase e da li-ase AP de eliminação β; 2) o C1 poderia converter o produto de eliminação de β para o produto de eliminação de δ; e 3) os sites AP formados na reação de glicosilase, poderiam servir como substrato para uma reação de eliminação de C1-catalítica β,δ. Para tratar essas possibilidades, foi gerado DNA duplex contendo um único site específico de PA através do tratamento do DNA contendo dU correspondente com a glicosilase de DNA uracil (UDG) (Fig. 1b) e reagiu com C1 ou hOGG1, seja individualmente ou em combinação. Tal como com o oligodeoxinucleótido 8-oxo-dG, a hOGG1 produziu o esperado produto β-eliminação, enquanto a adição de C1 produziu uma mistura de dois produtos (Fig. 1d). Esta mistura de produto também foi formada quando o oligodeoxinucleotídeo contendo AP foi incubado apenas com C1. Enquanto a posição da banda do produto de migração mais lenta correspondia ao produto de eliminação do hOGG1 β (Fig. 1d), a banda do produto de migração mais rápida co-migrou com o conhecido produto de eliminação do hNEIL1 do β (Fig. 1e). Assim, C1 promove as reações de eliminação β- e δ- nos sites AP.
A reação de clivagem por C1 no site AP contendo DNA foi dependente da concentração (Fig. 1e e Figura Complementar S1), sendo as baixas concentrações micromolares suficientes para observar os produtos. Semelhante às “nucleases artificiais “26 previamente desenhadas, C1 mostrou catálise de viragem no substrato de DNA: 1 pmol C1 gerou ~1,4 pmol produtos em 16 h (Fig. 1f). Assim, a taxa de incisão por C1 neste DNA foi de ~1,5 × 10-3 min-1 ou superior. A comparação de C1 com os reagentes AP lyase atualmente disponíveis espermina17 e o peptídeo KWKK23 demonstrou que o catalisador recém-identificado era pelo menos 100 vezes mais eficiente (Figura Suplementar S1).
Moieties estruturais transversais em C1 são importantes para a catálise
Análogos de C1 foram testados para determinar a contribuição de características estruturais específicas para a química da cisão dos fios. Inicialmente examinamos a capacidade das aminas simples, comercialmente disponíveis, que não possuem a fracção indolino-pirrol C2 (Fig. 2) e CS1-CS3 (Figura Complementar S2) de incisar o ADN num local AP. A formação de produtos de incisão, se houver, estava abaixo do nível de detecção em 10 μM. Assim, a meia indolinona-pirrol é importante para a reação. É possível que a subunidade indolinona-pirrol se ligue ao sítio AP29 ou ranhura menor e posicione a amina secundária para catalisar a clivagem do cordão através de um intermediário covalente com o sítio AP. Os compostos C3 (Fig. 2) e CS4 (Figura Complementar S2) que continham a fracção indolinona-pirrol mas não tinham a amina secundária também estavam completamente inativos. Estes dados sugerem fortemente o papel da amina como o grupo funcional reativo. A substituição do grupo metoxi na pirâmide por outras substituições, como em C4 e C5 (Fig. 2), levou a uma diminuição da atividade. A reação também foi modulada por um substituto no grupo amino, de modo que o aumento das demandas estéreis da amina secundária diminuiu a quantidade do produto observado (compare C1, C6 e C7 (Fig. 2) e C4, CS5 e CS6 (Figura Complementar S2)). Estas análises de estrutura-actividade demonstraram que várias das loites individuais são necessárias mas não suficientes para a clivagem, e que estas loites devem actuar cooperativamente para produzir a cisão dos fios.

(a) Estruturas de compostos representativos. (b) Substrato de DNA. (c) Ensaio para as capacidades dos compostos representativos (10 μM) para incisar o ADN (250 nM) no local de AP. Reações foram realizadas a 37 °C para 30 in.
A estrutura do DNA modula a clivagem C1-catalítica
A importância da moleza indolinona-pirrol para a química da cisão dos fios sugeriu que a estrutura do substrato de DNA poderia afetar a clivagem C1-mediada. Para abordar essas relações, as taxas iniciais de reação foram medidas para um DNA de cadeia única (a seqüência como na Fig. 1a) e os correspondentes DNAs de cadeia dupla que continham A, C, G, ou T em frente ao local da AP. O oligodeoxinucleotídeo homopolimérico, 5′-TAMRA-(T)5-AP-(T)11-3′, também foi examinado como uma estrutura de DNA de cadeia única inequívoca. Esses dados demonstraram que, embora a cisão dos fios pudesse ocorrer no contexto de DNA de cadeia única, os DNAs de cadeia dupla eram os substratos preferidos para o C1 (Fig. 3). Também foi constatado que a natureza da base oposta ao sítio AP modulava a catálise, sendo a taxa de hidrólise do sítio AP mais rápida em relação às pirimidinas opostas do que às purinas. Estas observações são consistentes com a proposta de que a subunidade indolinona-pirrolina interage com o DNA para ocupar um espaço vazio no local da PA. Notavelmente, a taxa inicial medida para o DNA de cadeia dupla com C oposto ao local da PA, foi muito próxima da taxa observada para este substrato sob condições de limitação da concentração de C1 (Fig. 1f).

Reações foram realizadas a 37 °C usando 250 nM de DNA e 5 μM C1. As taxas iniciais médias com os respectivos desvios padrão foram calculadas a partir de três experiências independentes usando o software KaleidaGraph 4.1 (Synergy Software). Os valores de P foram calculados usando o teste t de Student.
Cleavage of DNA at AP sites by C1 proceeds via an intermediate involving the secondary amine
To test for the covalent intermediate with the ring-opened, aldehydic form of the deoxyribose, an AP site-containing 32P-labeled oligodeoxynucleotide (Fig. 4a) foi incubado com C1 na presença de NaB(CN)H3. Enquanto este redutor reage lentamente com a lesão AP, ele prende eficientemente o imino- ou imino-conjugador de íons. Em uma reação de controle com o sítio AP contendo oligodeoxinucleotídeo e NaB(CN)H3 (Fig. 4b, pista 5), uma pequena fração (~3%) do DNA manifestou uma diminuição da mobilidade. Este produto de baixa abundância é comumente observado nas reações de aprisionamento23 e provavelmente representa um complexo com moléculas de Tris, como foi previamente demonstrado nas reações com o adutor de DNA de pirimidopurinona malondialdeído30. Em uma reação de controle positivo usando o peptídeo lisina-trriptofano-lisina-lisina (KWKK)23, o intermediário imina foi aprisionado, como evidenciado pela mudança na mobilidade do DNA (Fig. 4b, pista 3). Na presença de NaB(CN)H3 e C1, a maioria do DNA (~80%) formou um complexo que surgiu como uma espécie com mobilidade reduzida (Fig. 4b, pista 4). A formação do complexo não se deveu à ligação não específica de C1 ao DNA, uma vez que não foi observado nenhum deslocamento quando o oligodeoxinucleotídeo contendo dU correspondente foi testado sob condições idênticas (Fig. 4b, pista 1). Esses dados foram consistentes com a hipótese de que o reagente foi capaz de formar um íon intermediário de íon de imônio covalente, que então posiciona um grupo lateral para uma abstração de prótons do anel de açúcar.

(a) substratos de DNA. (b) Armadilha de cianoborohidreto de um complexo entre C1 e AP site contendo DNA. (c) Fragmentação do CID do íon iminio intermediário reduzido. (d) Fragmentação do conjugado C1-deoxirribose reduzido após digestão enzimática.
O aprisionamento redutor de C1 foi repetido usando oligodeoxinucleotídeo AP não rotulado contendo oligodeoxinucleotídeo (Fig. 4a), e o produto foi analisado por espectrometria de massa (EM). A análise revelou uma massa consistente com o complexo covalente de DNA-C1 reduzido (m/z 1134,64 para -2). A dissociação induzida por colisão (CID) deste íon resultou em um conjunto completo de íons a-B (Base) e w, consistente com o íon iminium intermediário reduzido entre o C1 e o local AP (Figura Complementar S3). Na série de íons a-B, a fragmentação da ligação C3′-O é normalmente acompanhada pela perda neutra da nucleobase. A ligação reduzida entre C1 e o sítio AP era esperada ser menos labial e como resultado, observamos o íon a4 (m/z 1330.1) bem como o íon a4-B (m/z 1005.6). O oligodeoxinucleotídeo também foi digerido enzimaticamente e analisado pela EM (Fig. 4c e Figura Suplementar S4). Foi observado um produto digestivo com uma massa consistente com o conjugado C1-deoxirribose reduzido (m/z 444,19); a fragmentação deste íon produto deu um íon filha com m/z 295,09, que resultou da perda neutra de N-metil amino-2-deoxirribitol (Fig. 4d e Figura S4 Suplementar). Este produto foi idêntico ao preparado a partir da reação de aminação redutora de 2-deoxirribose e C1. Estes estudos demonstram que o C1 cliva o DNA contendo AP através da catálise covalente envolvendo a amina secundária.
C1 aumenta a estabilidade térmica do DNA contendo um análogo de um sítio AP
O efeito do C1 foi estudado na estabilidade térmica do DNA contendo um tetrahidrofurano (THF), análogo estrutural de um sítio AP que é incapaz de sofrer a reação de eliminação31. O Tm do DNA duplex contendo THF oposto ao T (T:THF) aumentou 6,3 °C após a adição de 1 equivalente de C1, enquanto o Tm do DNA de controle contendo um par T:A foi menos afetado e só aumentou 1 °C (Fig. 5). Enquanto a curva de fusão de T:THF com C1 indicava estabilização térmica por C1, a presença de C1 também ampliou a curva de fusão, indicando uma transição de fusão menos cooperativa. Coletivamente, estas observações sugerem que C1 se liga especificamente ao DNA no local AP, fornecendo estabilização localizada à hélice de DNA. Isto é consistente com o modelo proposto de interações entre C1 e DNA, no qual a moleza indolinona-pirrol ocupa o espaço disponível no local AP.
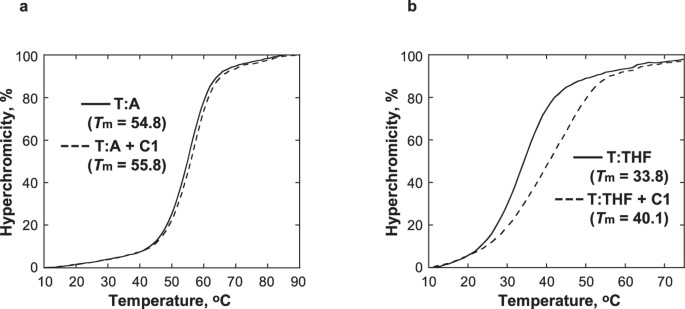
As curvas de temperatura de fusão de T:A (a) e T:THF (b) foram obtidas usando 5 μM DNA em tampão fosfato de sódio 10 mM, pH 7.0, 100 mM NaCl, e 1 mM EDTA na ausência (linha sólida) e presença (linha tracejada) de 5 μM C1.
C1 tem maior afinidade para DNA contendo um sítio AP
Para examinar o modo de ligação e afinidade de DNA contendo C1 para AP, análises de dicroísmo circular (CD) foram conduzidas usando ambos THF contendo e controle T:A duplex oligodeoxinucleotídeos (Fig. 6). O CD induzido (CDI) de C1 na interação com o duplex T:THF foi observado como um forte sinal de exciton, uma forma bisigna com bandas positivas e negativas em relação ao máximo de absorção do C1 livre. Isto é geralmente indicativo da formação de complexos diméricos ou de ordem superior, seja em modo de encadernação de ranhuras ou em modo de empilhamento externo32. A titulação de C1 com concentrações crescentes de T:THF (Fig. 6a e Figura Complementar S5) revelou tanto interações não específicas como específicas. No início da titulação, C1 está em excesso e as interações não-específicas são favorecidas. Com a adição de T:THF, o CDI a ~488 nm aumentou. Em aproximadamente 2 μM concentração de DNA, a banda de CDI mudou para uma energia mais baixa (~495 nm) e depois diminuiu até o ponto de equivalência e platô em excesso de DNA indicando uma interação específica. A titulação de C1 com o DNA T:A de controle (Fig. 6a e Figura Complementar S5) não apresentou o CDI positivo em torno de 495 nm, indicando a ausência de interações específicas. A titulação de T:THF também exibiu evidência de interações não específicas e específicas (Fig. 6b). Em baixas concentrações de C1 com DNA em excesso, a banda de CDI foi centralizada em ~495 nm (específico); como a concentração de C1 aumentou em relação ao DNA, a intensidade do CDI aumentou e mudou para uma energia mais alta (~488 nm), indicativo de ligação não específica como esperado em altas concentrações de ligante. O deslocamento hipocrômico observado pode indicar uma alteração ou uma conformação mais definida de C1 quando especificamente ligado ao local da PA. A titulação do DNA de controle sem um local de PA também mostrou um CDI e evidência de ligação não específica (Fig. 6c). Vale notar que a faixa de 450-520 nm só apareceu com um excesso de 3 vezes de C1 em relação ao DNA, tornando esta faixa de comprimento de onda a melhor escolha para construir as curvas de ligação. Isotermas de ligação foram construídas e analisadas usando um modelo simples de ligação bimolecular, conforme publicado anteriormente32. As equações não lineares resultantes das isotermas de 500 nm (Fig. 6d) deram a constante de dissociação (KD) de 64 μM para o DNA de controle T:A (ligação não específica) e 29 μM para o DNA T:THF (ligação específica); o KD de C1 com duplex contendo THF foi calculado para ser 22 μM após subtração para corrigir a contribuição de ligação não específica e possíveis interações ligante-ligante (Tabela 1). Os valores de KD calculados a 495 nm estavam apenas ligeiramente fora dos erros dos calculados a 500 nm (Tabela 1).

(a) Curvas de titulação de intensidade de CDI a 500 nm usando uma concentração constante de C1 (20 μM) e concentrações crescentes de DNA (titulação reversa). (b) Espectros de CD para T:THF e (c) T:A usando uma concentração constante de DNA (10 μM) e concentrações crescentes de C1 (titulação progressiva). (d) Curvas de titulação de intensidade de CDI a 500 nm versus o logaritmo da concentração de C1 para T:THF e T:A (derivadas dos dados nos painéis b e c). (e) Curvas de titulação da intensidade do CDI a 500 nm versus o logaritmo da concentração de C6 para T:THF e T:A (derivadas dos dados da Figura Complementar S6). (d) Curvas de titulação da intensidade do CDI a 500 nm versus o logaritmo da concentração de C6 para T:AP e T:A (derivadas dos dados da Figura Suplementar S6).
Outra, foi considerada a possibilidade de que a afinidade de C1 poderia ser diferente para o DNA contendo um sítio natural de AP em vez de THF. Para abordar esta questão, nós testamos o composto C6 estruturalmente relacionado mas inativo (Fig. 2) em análises de CD com DNA contendo THF ou um local de AP que foi criado a partir do tratamento dU por UDG. Os espectros de CD observados para o C6 em experimentos de titulação dianteira usando o T:THF e T:A DNAs (Figura Complementar S6) foram geralmente similares aos observados para o C1 (Fig. 6b,c). As isotermas de ligação foram construídas (Fig. 6e) e analisadas como acima. O KD de C6 com duplex contendo THF, corrigido para as interações não específicas foi calculado para ser 28 μM (Tabela 1), que é apenas ~25% maior que o KD correspondente calculado para C1. Assim, C6 parece ser um modelo apropriado para estudar as interações de C1 com o DNA. A titulação de C6 foi então realizada usando o DNA que continha o sítio AP derivado da UDG (Figura Complementar S6). A análise das isotermas de ligação revelou que a afinidade de C6 com este ADN era essencialmente idêntica ao ADN contendo THF (Fig. 6f e Tabela 1). Considerando que a desoxirribose em um local de PA natural existe predominantemente na forma de anel fechado, semelhante ao THF31, o último resultado não foi surpreendente. Pode-se prever que antes da formação de um intermediário covalente, a afinidade do ADN C1 com o AP seria comparável à medida para o ADN contendo THF. Assim, a ligação mais forte de C1 ao DNA contendo um local AP em comparação com o DNA de controle foi consistente com os dados de estabilidade térmica e, em conjunto, apoia a proposta de que a subunidade indolinona-pirrol ocupa um espaço vazio no local AP do DNA. É provável que a afinidade de C1 para o local AP contribua para sua atividade lásica e forneça uma clara vantagem sobre C2, CS1, CS2 e CS3, que carecem da fracção indolinona-pirrol (Fig. 2 e Figura Complementar S2).
Conclusões
Os catalisadores de cisão de cadeia de DNA AP descritos neste estudo representam uma nova classe de compostos que podem ser utilizados para clivar locais AP sob condições fisiológicas. Com base nesta estrutura central, as moléculas podem ser projetadas para melhorar a seletividade da incisão do local AP em bases opostas e potencialmente para contextos sequenciais diferentes. Devido ao recente progresso no desenvolvimento de técnicas de triagem de alto rendimento, estes compostos podem ser rapidamente avançados para serem reagentes eficientes na clivagem de locais de PA que são criados através da depuração ou da via de BER em células e organismos. As versões otimizadas de C1 também podem ter aplicações terapêuticas, particularmente em combinação com muitos medicamentos anticancerígenos comuns que danificam o DNA, como os agentes alquilantes33 , ou reparam o DNA alvo, como os inibidores PARP34,35 ou AP endonuclease36. Vale mencionar que o 3′ termina criado via β- ou β,3′- a eliminação não pode ser utilizada por polimerases de DNA sem reparo prévio37. As conseqüências biológicas da conversão de locais de PA em tais quebras de fios de DNA devem ser complexas e variar dependendo da capacidade celular de reparar ou tolerar diferentes tipos de danos de DNA. Os possíveis resultados podem incluir um aumento da eficácia terapêutica dos tratamentos anti-cancerígenos (morte celular mais eficiente) e uma diminuição da mutagénese induzida por drogas. A clivagem do DNA em locais de PA pode ser particularmente benéfica para o tratamento de cânceres que têm defeitos nos mecanismos de reparação de quebras de fios de DNA, tais como cânceres deficientes de BRCA34,35,