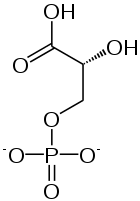
1,3-bisfosfoglicerato é a base conjugada do ácido 1,3-bisfosfoglicérico. É fosforilado no número 1 e 3 carbonos. O resultado desta fosforilação dá a 1,3BPG propriedades biológicas importantes, como a capacidade de fosforilato ADP para formar a molécula de armazenamento de energia ATP.
Em glycolysisEdit
| D-glyceraldehydehyde 3-phosphate | glyceraldehyde phosphate dehydrogenase | 1,3-bisfosfo-D-glicerato | 3-fosfoglicerato kinase | 3-fosfo-D-glicerato | ||
 |
 |
 |
||||
| NAD+ + Pi |
NADH + H+ |
ADP | ATP | |||
|
|
|
|||||
| NAD+ + Pi |
NADH + H+ |
ADP | ATP | |||
| gliceraldeído fosfato desidrogenase | 3-phosphoglycerate kinase | |||||
Compound C00118 no KEGG Pathway Database. Enzima 1.2.1.12 na base de dados de caminhos do KEGG. Composto C00236 na base de dados KEGG Pathway Database. Enzima 2.7.2.3 na base de dados de caminhos do KEGG. Composto C00197 no KEGG Pathway Database.
Como mencionado anteriormente 1,3BPG é um intermediário metabólico na via glicolítica. Ele é criado pela oxidação exergônica do aldeído em G3P. O resultado desta oxidação é a conversão do grupo aldeído em um grupo ácido carboxílico que impulsiona a formação de uma ligação de fosfato de acilo. Este é, aliás, o único passo na via glicolítica em que o NAD+ é convertido em NADH. A reação de formação da 1,3BPG requer a presença de uma enzima chamada gliceraldeído-3-fosfato desidrogenase.
A ligação de fosfato de acilo de alta energia de 1,3BPG é importante na respiração, pois auxilia na formação de ATP. A molécula de ATP criada durante a reação seguinte é a primeira molécula produzida durante a respiração. A reacção ocorre da seguinte forma;
1,3-bisfosfoglicerato + ADP ⇌ 3-fosfoglicerato + ATP
A transferência de um fosfato inorgânico do grupo carboxilo em 1,3BPG para ADP para formar ATP é reversível devido a uma baixa ΔG. Isto é como resultado de uma ligação de fosfato acilo ser clivada enquanto outra é criada. Esta reação não é naturalmente espontânea e requer a presença de um catalisador. Este papel é desempenhado pela enzima fosfoglicérato quinase. Durante a reação o fosfoglicérato quinase sofre uma mudança conformacional induzida pelo substrato semelhante a outra enzima metabólica chamada hexoquinase.
Porque duas moléculas de glicolida-3-fosfato são formadas durante a glicólise a partir de uma molécula de glicose, pode-se dizer que 1,3BPG é responsável por duas das dez moléculas de ATP produzidas durante todo o processo. A glicólise também utiliza duas moléculas de ATP em seus estágios iniciais como uma etapa comprometida e irreversível. Por este motivo, a glicólise não é reversível e tem uma produção líquida de 2 moléculas de ATP e duas de NADH. As duas moléculas de NADH em si vão produzir aproximadamente 3 moléculas de ATP cada.
Clique nos genes, proteínas e metabólitos abaixo para ligar aos respectivos artigos.

|{{{{bSize}}}px|alt=Glycolysis e Gluconeogenesis editar]]
- ^ O mapa interactivo de percursos pode ser editado no WikiPathways: “GlycolysisGluconeogenesis_WP534”.
No ciclo CalvinEdit
1,3-BPG tem um papel muito semelhante no ciclo Calvin ao seu papel na via glicolítica. Por esta razão, ambas as reacções são ditas análogas. No entanto, a via de reação é efetivamente invertida. A única outra grande diferença entre as duas reações é que o NADPH é usado como doador de elétrons no ciclo de Calvin, enquanto o NAD+ é usado como um aceitador de elétrons na glicólise. Neste ciclo de reação 1,3BPG origina-se do 3-fosfoglicérato e é transformado em 3-fosfato de glicerol pela ação de enzimas específicas.
Contrário às reações similares da via glicolítica, 1,3BPG no ciclo de calvina não produz ATP, mas sim o utiliza. Por este motivo pode ser considerado uma etapa irreversível e comprometida no ciclo. O resultado desta seção do ciclo é que um fosfato inorgânico é removido do 1,3BPG como um íon de hidrogênio e dois elétrons são adicionados ao composto+.
No completo reverso da reação da via glicolítica, a enzima fosfoglicérato quinase catalisa a redução do grupo carboxil de 1,3BPG para formar um aldeído em seu lugar. Esta reação também libera uma molécula inorgânica de fosfato que é posteriormente utilizada como energia para a doação de elétrons a partir da conversão do NADPH para NADP+. Supervisionando este último estágio da reação está a enzima gliceraldeído-fosfato desidrogenase.
Em transferência de oxigênioEditar
Durante o metabolismo normal em humanos aproximadamente 20% do 1,3BPG produzido não vai mais longe na via glicolítica. Em vez disso, é desviado por um caminho alternativo envolvendo a redução do ATP nos eritrócitos. Durante esta via alternativa é transformada numa molécula semelhante chamada ácido 2,3-bisfosfoglicérico (2,3BPG). O 2,3BPG é usado como mecanismo para supervisionar a liberação eficiente de oxigênio da hemoglobina. Os níveis deste 1,3BPG aumentarão no sangue do paciente quando os níveis de oxigênio estiverem baixos, pois este é um dos mecanismos de aclimatação. Níveis baixos de oxigênio provocam um aumento nos níveis de 1,3BPG que, por sua vez, eleva o nível de 2,3BPG que altera a eficiência da dissociação do oxigênio da hemoglobina.