Introducción
He tenido la suerte de participar en dos grandes expediciones de investigación fisiológica a altitudes extremas. La primera fue la expedición Silver Hut en 1960-1961, durante la cual un grupo de fisiólogos pasó varios meses a la altísima altitud de 5.800 m, y medimos los cambios fisiológicos que se produjeron durante este largo período. Se realizaron estudios adicionales hasta los 7.830 m. (25.700 pies). El objetivo general era determinar los mecanismos por los que las personas que normalmente viven cerca del nivel del mar responden a la hipoxia severa durante un período prolongado.
La segunda expedición fue la Expedición Americana de Investigación Médica al Everest que tuvo lugar en 1981. Aquí el objetivo fisiológico era muy diferente. El objetivo era obtener los primeros datos fisiológicos humanos en la cima del monte Everest (8.848 m, 29.028 pies) para aclarar cómo las personas que normalmente viven a nivel del mar pueden sobrevivir a la hipoxia extrema del punto más alto del mundo.
Expedición Silver Hut
Esta fue la idea de Sir Edmund Hillary que, junto con Tenzing Norgay, fue la primera persona en alcanzar la cima del monte Everest siete años antes. Hillary había colaborado con Griffith Pugh, un fisiólogo de gran altitud, en esta primera ascensión al Everest, y ambos estaban intensamente interesados en el proceso de aclimatación que permite a las personas que se encuentran cerca del nivel del mar ascender a altitudes muy elevadas.
Mi interés particular era la capacidad de difusión del pulmón. Barcroft (1925) había sugerido que el ejercicio a gran altura provocaría hipoxemia arterial debido a la limitación de la difusión a través de la barrera de gas en la sangre. Para comprobarlo, hicimos que los miembros de la expedición trabajaran a su máxima capacidad en un ergómetro de bicicleta, y medimos la saturación arterial de oxígeno utilizando un oxímetro de oído recientemente disponible (West et al., 1962). Descubrimos que, de hecho, había una marcada caída de la saturación arterial de oxígeno a pesar del aumento de la PO2 alveolar a medida que aumentaba el nivel de trabajo. Esto era una fuerte evidencia de la limitación de la difusión en estas condiciones de hipoxia severa. Las mediciones duplicadas en mí mismo mostraron una saturación de oxígeno de sólo el 33% en el ejercicio máximo, lo que reflejaba una hipoxemia muy severa.
También medimos la capacidad de difusión del monóxido de carbono en el transcurso de la expedición y mostramos que ésta apenas cambió (West, 1962). El pequeño aumento podría explicarse por la policitemia que se desarrolló. La conclusión fue que las características de la barrera de gases de la sangre no se alteraban por la exposición prolongada a la hipoxia severa. Esta fue la primera demostración clara de la limitación de difusión de la transferencia de oxígeno por el pulmón durante el ejercicio severo a gran altura.
Más tarde en la expedición, se hicieron mediciones de la captación máxima de oxígeno durante el ejercicio a la altitud extremadamente alta de 7.440 m (24.400 pies) (Pugh et al., 1964). La extrapolación de estos datos a la altitud de la cumbre del Everest sugirió que sería imposible alcanzar la cumbre sin oxígeno suplementario. Los estudios sanguíneos mostraron una marcada policitemia en los sujetos que vivían a una altitud de 5.800 m. Las concentraciones medias de hemoglobina y los hematocritos eran de 19,6 g/dl y 55,8%, respectivamente. Había pruebas de que el aumento inicial del hematocrito se debía principalmente a la pérdida de volumen plasmático, pero posteriormente se producía un gran aumento de la masa de glóbulos rojos. El electrocardiograma de las personas que vivían a 5.800 m mostraba una marcada hipertrofia ventricular derecha, y en algunos trazados había inversión de las ondas T en las derivaciones torácicas, lo que presumiblemente indicaba una hipoxia miocárdica grave (Milledge, 1963). Se realizaron mediciones de la función neuropsicométrica mediante la clasificación de tarjetas, y se descubrió que la tasa de clasificación estaba reducida, pero que con el aumento de la concentración los sujetos podían clasificar sin errores. Todos los miembros de la expedición sufrieron una severa e incesante pérdida de peso mientras vivían a 5.800 m, a un ritmo de entre 0,5 y 1,5 kg/semana. La impresión general era que no sería posible que las personas vivieran indefinidamente a esta gran altitud (Pugh, 1962).
Expedición Americana de Investigación Médica al Everest
Como se ha señalado anteriormente, el objetivo de esta expedición era aclarar cómo los seres humanos pueden tolerar la hipoxia de la mayor altitud del mundo. Sorprendentemente, unos meses antes de que tuviera lugar la expedición, dos alpinistas europeos alcanzaron por primera vez la cima del Monte Everest sin utilizar oxígeno suplementario. Esta hazaña asombró a muchos fisiólogos y suscitó muchas preguntas sobre cómo pudo hacerse.
El programa de investigación fue muy extenso, y aquí sólo se puede dar un breve resumen. Se realizaron mediciones en el campamento base, a 5.400 m de altitud, y en el campamento base avanzado, a 6.300 m de altitud, y en el campamento más alto, a 8.050 m. También esperábamos conseguir algunas mediciones en la cumbre del Everest, aunque esto era muy ambicioso. De hecho, cuando miramos hacia atrás en las seis expediciones anteriores a la nuestra, ninguna de ellas llegó a la cumbre. Si el tiempo es malo, olvídalo, y un factor crítico es si los suficientes miembros de la expedición se mantienen en forma a pesar de la extrema altitud.
En el campamento base, medimos la respuesta ventilatoria a la hipoxia, es decir, la medida en que la respiración aumenta cuando el sujeto se expone a una mezcla de oxígeno inspirado baja. Los resultados fueron sorprendentes. Resultó que el escalador que llegó primero a la cumbre tuvo la respuesta más alta, el que llegó segundo a la cumbre tuvo la siguiente respuesta más alta, y el escalador que llegó tercero a la cumbre tuvo la tercera respuesta más alta (Schoene et al., 1984). Esto debe ser en parte una casualidad, pero sin duda sugiere que existe una relación entre la medida en que los escaladores aumentan su ventilación y su tolerancia a la altitud extrema. La razón de esto se aclarará más adelante.
Se llevaron a cabo un gran número de estudios en el campo base avanzado, pero sólo uno, un estudio neuropsicométrico, se resumirá aquí. Es bien sabido que el cerebro y el sistema nervioso central son muy sensibles a la hipoxia. Por ejemplo, si alguien se cae en una piscina y es rescatado 5 o 10 minutos después, puede ser reanimado con éxito, pero el sistema nervioso central nunca se recupera del todo. Por lo tanto, no fue sorprendente que pudiéramos mostrar cambios en mediciones como la memoria a corto plazo y la habilidad manipulativa (determinada a partir de una prueba de golpeo con el dedo) en las altitudes muy elevadas. Esto no era inesperado. Sin embargo, cuando la expedición regresó al nivel del mar, se comprobó que dos de las mediciones seguían siendo anormales. Se trataba de la memoria a corto plazo y de la prueba de golpeteo con el dedo (Townes et al., 1984). Por lo tanto, estaba claro que cualquier persona que regrese de estas altitudes extremas es probable que tenga algún deterioro residual del sistema nervioso central. Fuimos uno de los primeros grupos en demostrarlo, pero se ha confirmado muchas veces desde entonces.
Algunos de los hallazgos más interesantes fueron los de la cumbre. Habíamos diseñado y construido un dispositivo especial que permitía al escalador recoger el último gas expirado tras una espiración máxima, es decir, una muestra de gas alveolar. Más de 34 muestras, incluidas cuatro de la cima, se llevaron a la UC San Diego en botes herméticos al gas. Cuando se comparó la PCO2 alveolar con la presión barométrica, que disminuía a medida que aumentaba la altitud, se descubrió que la PCO2 en la cumbre era de 7-8 mm Hg. Se trataba de un valor casi increíblemente bajo, ya que el valor a nivel del mar es de unos 40 mm Hg. Este valor extremadamente bajo subraya el enorme aumento de la ventilación alveolar que es necesario en estas altitudes extremas (West et al., 1983).
Cuando se trazaron tanto la PO2 como la PCO2 alveolares frente a la presión barométrica, surgió una imagen interesante (Figura 1). Tanto la PO2 como la PCO2 disminuyeron a medida que aumentaba la altitud. La caída de la PO2 se produjo por la reducción del aire alrededor del escalador como resultado de la reducción de la presión barométrica. El descenso de la PCO2 fue causado únicamente por la hiperventilación del escalador. Resultó que cuando la altitud superó los 7000 m aproximadamente, no hubo más caída de la PCO2 alveolar. La figura muestra que ésta se defiende a un nivel de unos 35 mm Hg. En otras palabras, para sobrevivir a estas enormes altitudes, es necesario montar una ventilación alveolar que haga descender la PCO2 a unos 8 mmHg y así preservar la PO2 alveolar en el nivel muy bajo pero viable de unos 35 mm Hg. Esto explica por qué en las mediciones de la respuesta ventilatoria a la hipoxia a las que se ha hecho referencia anteriormente, había una correlación entre la magnitud de la respuesta y la tolerancia del alpinista a la altitud extrema. Si no es capaz de montar un grado de hiperventilación que sea suficiente para hacer descender la PCO2 alveolar a unos 7-8 mm Hg, no podrá mantener un nivel viable de PO2 en el gas alveolar. Así pues, la hiperventilación extrema es una de las características más importantes de la respuesta fisiológica a la altitud extremadamente elevada.
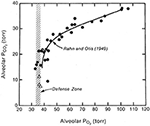
Figura 1. Valores de la PO2 y la PCO2 en el gas alveolar de los alpinistas a medida que ascienden desde el nivel del mar (arriba a la derecha) hasta la cumbre del Everest (abajo a la izquierda). La PO2 disminuye debido a la reducción de la presión barométrica. La PCO2 disminuye debido al aumento de la ventilación alveolar. Por encima de una altitud correspondiente a una PCO2 de unos 20 mm Hg (unos 7000 m), no hay más caída de la PO2. En otras palabras, ésta se defiende a unos 35 mmHg. Esto sólo puede hacerse si la PCO2 se reduce continuamente más por la hiperventilación extrema a medida que aumenta la altitud. Modificado de Rahn y Otis (1949) y West et al. (1983).
La PCO2 alveolar extremadamente baja suscita la pregunta de qué ocurrió con el pH arterial. Es razonable suponer que la PCO2 arterial y la alveolar son las mismas. Afortunadamente, dos de los alpinistas se tomaron sangre venosa mutuamente a la mañana siguiente de su exitoso ascenso a la cumbre, por lo que se pudieron medir los valores de exceso de base. Cuando estos valores se introdujeron en la ecuación de Henderson-Hasselbalch, el pH arterial resultante estaba entre 7,7 y 7,8. Este es un grado extremo de alcalosis respiratoria.
Una pregunta interesante es por qué el riñón no eliminó bicarbonato para desarrollar una compensación metabólica para esta alcalosis extrema y así mover el pH más cerca de lo normal. Esta es la respuesta habitual si se genera una alcalosis respiratoria, por ejemplo, por hiperventilación como ocurre a veces durante la histeria. La razón de la ausencia de compensación metabólica no se entiende completamente, pero una posibilidad es que estos escaladores estaban probablemente muy agotados de volumen. Esta es una característica común de ir a gran altitud y, por ejemplo, los escaladores que se quedaron en el campamento base avanzado a 6300 m mostraron evidencia de agotamiento crónico de volumen. Un factor responsable en la altitud extrema es presumiblemente la enorme hiperventilación, pero la sed también se ve afectada.
Las consecuencias fisiológicas de la alcalosis severa son interesantes. Otros estudios han demostrado que una mayor afinidad de la hemoglobina por el oxígeno aumenta la supervivencia en un entorno hipóxico. Hace muchos años, se demostró que mamíferos como la vicuña y la llama en los Andes sudamericanos tienen una mayor afinidad por el oxígeno (es decir, tienen una curva de disociación de oxígeno desplazada hacia la izquierda) en comparación con los mamíferos que viven a nivel del mar (Hall, 1937). Así, los alpinistas a gran altura tienen la misma respuesta.
También es cierto que si se observa de forma general en todo el reino animal a los organismos que viven en un ambiente hipóxico, muchos de ellos han desarrollado una mayor afinidad de la hemoglobina por el oxígeno. Uno de los mejores ejemplos es el feto humano que, debido a una diferencia en la secuencia de aminoácidos de la hemoglobina, tiene un marcado aumento de la afinidad por el oxígeno con una P50 de unos 19 mm Hg comparada con la de unos 27 para un adulto. El feto humano presenta una hipoxemia severa según los estándares de los adultos, con una PO2 en la aorta descendente de unos 22 mm Hg, que es incluso inferior a la de un escalador en la cumbre del Everest. Es fascinante que el alpinista exitoso tenga la ventaja de una mayor afinidad de la hemoglobina por el oxígeno. Esto ayuda a la carga de oxígeno en el capilar pulmonar. Se podría argumentar que también interfiere con la descarga de oxígeno en la periferia del cuerpo, pero los estudios han demostrado que la ventaja de la carga en el pulmón es mayor que la desventaja en la descarga en los tejidos periféricos.
Una pregunta interesante es cuál es el consumo máximo de oxígeno de un escalador en la cumbre. Como se ha indicado anteriormente, las mediciones previas realizadas a una altitud extrema durante la expedición de Silver Hut sugerían que todo el oxígeno disponible en la cumbre sería necesario para la actualización del oxígeno basal, es decir, para mantener el bombeo del corazón y la actividad del cerebro. Era imposible colocar un ergómetro para bicicletas en la cumbre. Sin embargo, tomamos a escaladores que estaban muy bien aclimatados y les hicimos pedalear al máximo a una altitud de 6300 m mientras respiraban un 14% de oxígeno. Esto les dio la misma PO2 inspirada que en la cumbre. El nivel de consumo de oxígeno en estas condiciones fue de aproximadamente 1 L/min, lo cual es un consumo máximo de oxígeno miserablemente bajo, y es equivalente a alguien que camina lentamente en el nivel. Sin embargo, aparentemente es suficiente para permitir que un escalador llegue a la cumbre.
Contribuciones de los autores
JW redactó el manuscrito, leyó y aprobó la versión presentada.
Declaración de conflicto de intereses
El autor declara que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un potencial conflicto de intereses.
Barcroft, J. (1925). The Respiratory Function of the Blood. Part 1 Lessons from High Altitudes. Cambridge: Cambridge University Press.
Google Scholar
Hall, F. G. (1937). Adaptaciones de los animales a la altura. J. Mammol. 18, 469-472. doi: 10.2307/1374337
CrossRef Full Text | Google Scholar
Milledge, J. S. (1963). Electrocardiographic changes at high altitude. Br. Heart J. 15, 291-298. doi: 10.1136/hrt.25.3.291
CrossRef Full Text | Google Scholar
Pugh, L. G. (1962). Aspectos fisiológicos y médicos de la Expedición Científica y del Himalaya 1960-61. Br. Med. J. 2, 621-633. doi: 10.1136/bmj.2.5305.621
CrossRef Full Text | Google Scholar
Pugh, L. G., Gill, M. B., Lahiri, S., Milledge, J. S., Ward, M. P., y West, J. B. (1964). Muscular exercise at great altitudes. J. Appl. Physiol 19, 431-440. doi: 10.1152/jappl.1964.19.3.431
CrossRef Full Text | Google Scholar
Rahn, H., y Otis, A. B. (1949). La respuesta respiratoria del hombre durante y después de la aclimatación a gran altura. Am. J. Physiol. 157, 445-462. doi: 10.1152/ajplegacy.1949.157.3.445
PubMed Abstract | CrossRef Full Text | Google Scholar
Schoene, R. B., Lahiri, S., Hackett, P. H., Peters, R. M., Milledge, J. S., Pizzo, C. J., et al. (1984). Relationship of hypoxic ventilatory response to exercise performance on Mt Everest. J. Appl. Physiol. 56, 1478-1483. doi: 10.1152/jappl.1984.56.6.1478
CrossRef Full Text | Google Scholar
Townes, B. D., Hornbein, T. H., y Grant, I. (1984). «Human cerebral function at extreme altitude», en High Altitude and Man, eds J. B. West y S. Lahiri (Bethesda, MD: American Physiological Society), 31-36.
Google Scholar
West, J. B. (1962). Capacidad de difusión del pulmón para el monóxido de carbono a gran altura. J. Appl. Physiol. 17, 421-426. doi: 10.1152/jappl.1962.17.3.421
CrossRef Full Text | Google Scholar
West, J. B., Hackett, P. H., Maret, K. H., Milledge, J. S., Peters, R. M., Pizzo, C. J., et al. (1983). Pulmonary gas exchange on the summit of Mt Everest. J. Appl. Physiol. 55:678-687. doi: 10.1152/jappl.1983.55.3.678
CrossRef Full Text | Google Scholar
West, J. B., Lahiri, S., Gill, M. B., Milledge, J. S., Pugh, L. G. C. E., and Ward, M. P. (1962). Arterial oxygenation during exercise at high altitude. J. Appl. Physiol. 17, 617-621. doi: 10.1152/jappl.1962.17.4.617
CrossRef Full Text | Google Scholar