El destino metabólico (catabólico) del propionil-CoA depende del entorno en el que se sintetice. Por lo tanto, el propionil-CoA en un ambiente anaeróbico podría tener un destino diferente que en un organismo aeróbico. Las múltiples vías, ya sea el catabolismo por la propionil-CoA carboxilasa o la metilcitrato sintasa, también dependen de la presencia de varios genes.
- Reacción con la propionil-CoA carboxilasaEditar
- MecanismoEditar
- Ciclo del metilcitratoEditar
- Metabolismo bacterianoEditar
- Metabolismo de Mycobacterium tuberculosisEditar
- Posible secuestro en R. sphaeroidesEditar
- Metabolismo de Escherichia coliEditar
- Metabolismo de las plantasEditar
- Metabolismo de los hongosEditar
- Propionilación de proteínasEditar
Reacción con la propionil-CoA carboxilasaEditar
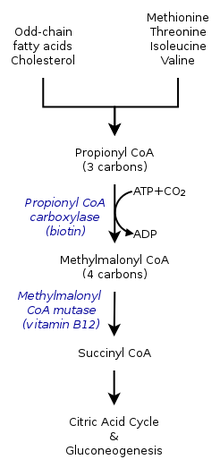
Dentro del ciclo del ácido cítrico en los seres humanos, el propionil-CoA, que interactúa con el oxaloacetato para formar metilcitrato, también puede catalizarse en metilmalonil-CoA a través de la carboxilación por la propionil-CoA carboxilasa (PCC). El metilmalonil-CoA se transforma posteriormente en succinil-CoA para su posterior utilización en el ciclo del ácido tricarboxílico. La PCC no sólo cataliza la carboxilación del propionil-CoA a metilmalonil-CoA, sino que también actúa sobre varios acil-CoAs diferentes. Sin embargo, su mayor afinidad de unión es con el propionil-CoA. Además, se demostró que la transformación del propionil-CoA se inhibe en ausencia de varios marcadores del TCA, como el glutamato. El mecanismo se muestra en la figura de la izquierda.
MecanismoEditar
En los mamíferos, el propionil-CoA se convierte en (S)-metilmalonil-CoA por la propionil-CoA carboxilasa, una enzima dependiente de la biotina que también requiere bicarbonato y ATP.
Este producto se convierte en (R)-metilmalonil-CoA por la metilmalonil-CoA racemasa.
(R)-metilmalonil-CoA se convierte en succinil-CoA, un intermediario en el ciclo del ácido tricarboxílico, por la metilmalonil-CoA mutasa, una enzima que requiere

cobalamina para catalizar la migración del enlace carbono-carbono.
El mecanismo de la metilmalonil-CoA mutasa comienza con la escisión del enlace entre el 5′ CH
2- del 5′-desoxiadenosilo y el cobalto, que está en su estado de oxidación 3+ (III), lo que produce un radical de 5′-desoxiadenosilo y cobalamina en el estado de oxidación reducido de Co(II).
A continuación, este radical abstrae un átomo de hidrógeno del grupo metilo de la metilmalonil-CoA, lo que genera un radical metilmalonil-CoA. Se cree que este radical forma un enlace carbono-cobalto con la coenzima, al que sigue la reordenación del esqueleto de carbono del sustrato, produciendo así un radical succinil-CoA. Este radical pasa a abstraer un hidrógeno de la 5′-desoxiadenosina producida anteriormente, creando de nuevo un radical desoxiadenosilo, que ataca a la coenzima para reformar el complejo inicial.
Un defecto en la enzima metilmalonil-CoA mutasa da lugar a la aciduria metilmalónica, un peligroso trastorno que provoca una disminución del pH sanguíneo.

Ciclo del metilcitratoEditar
La acumulación de propionil-CoA puede resultar tóxica para diferentes organismos. Dado que se han propuesto diferentes ciclos sobre cómo se transforma el propionil-CoA en piruvato, un mecanismo estudiado es el ciclo del metilcitrato.La reacción inicial es la beta-oxidación para formar el propionil-CoA que se descompone posteriormente por el ciclo. En esta vía intervienen tanto las enzimas relacionadas con el ciclo del metilcitrato como las del ciclo del ácido cítrico. Todas ellas contribuyen a la reacción global de desintoxicación de las bacterias de la dañina propionil-CoA. También se atribuye como vía resultante debido al catabolismo de los ácidos grasos en las micobacterias. Para proceder, el gen prpC codifica la metilcitrato sintasa, y si no está presente, el ciclo del metilcitrato no se producirá. En su lugar, el catabolismo procede a través de la propionil-CoA carboxilasa. Este mecanismo se muestra abajo a la izquierda junto con los reactantes, productos, intermedios y enzimas participantes.
Metabolismo bacterianoEditar
Metabolismo de Mycobacterium tuberculosisEditar
La oxidación de propionil-CoA para formar piruvato está influenciada por su necesidad en Mycobacterium tuberculosis. La acumulación de propionil-CoA puede dar lugar a efectos tóxicos. En Mycobacterium tuberculosis, se ha sugerido que el metabolismo del propionil-CoA está implicado en la biogénesis de la pared celular. Por lo tanto, la falta de dicho catabolismo aumentaría la susceptibilidad de la célula a diversas toxinas, especialmente a los mecanismos antimicrobianos de los macrófagos. Otra hipótesis sobre el destino del propionil-CoA, en M. tuberculosisis, es que, dado que el propionil-CoA se produce por el catabolismo de los ácidos grasos de cadena impar, el ciclo del metilcitrato se activa posteriormente para anular cualquier toxicidad potencial, actuando como mecanismo de amortiguación.
Posible secuestro en R. sphaeroidesEditar
El propionil-CoA tiene puede tener muchos efectos adversos y tóxicos en diferentes especies, incluida la bacteria. Por ejemplo, la inhibición de la piruvato deshidrogenasa por una acumulación de propionil-CoA en Rhodobacter sphaeroides puede resultar mortal. Además, al igual que en el caso de E. coli, una afluencia de propionil-CoA en especies de Miobacterias puede provocar toxicidad si no se trata inmediatamente. Esta toxicidad está causada por una vía que implica a los lípidos que forman la pared celular bacteriana. Mediante la esterificación de los ácidos grasos de cadena larga, el exceso de propionil-CoA puede ser secuestrado y almacenado en el lípido triacilglicerol (TAG), lo que conduce a la regulación de los niveles elevados de propionil-CoA. Este proceso de ramificación metílica de los ácidos grasos hace que actúen como sumideros para la acumulación de propión
Metabolismo de Escherichia coliEditar
En una investigación realizada por Luo et al., se utilizaron cepas de Escherichia coli para examinar cómo el metabolismo del propionil-CoA podría conducir potencialmente a la producción de ácido 3-hidroxipropiónico (3-HP). Se demostró que una mutación en un gen clave implicado en la vía, la succinato CoA-transferasa, conducía a un aumento significativo de 3-HP. Sin embargo, éste es todavía un campo en desarrollo y la información sobre este tema es limitada.

Metabolismo de las plantasEditar
El metabolismo de los aminoácidos en las plantas se ha considerado un tema controvertido, debido a la falta de pruebas concretas de alguna vía en particular. Sin embargo, se ha sugerido que están implicadas las enzimas relacionadas con la producción y uso de propionil-CoA. Asociado a esto está el metabolismo del isobutiril-CoA. Estas dos moléculas se consideran intermedias en el metabolismo de la valina. Como el propionato se presenta en forma de propionil-CoA, se descubrió que el propionil-CoA se convierte en β-hidroxipropionato a través de una vía de β-oxidación enzimática peroxisomal. Sin embargo, en la planta Arabidopsis no se observaron enzimas clave en la conversión de valina en propionil-CoA. A través de diferentes experimentos realizados por Lucas et al., se ha sugerido que en las plantas, a través de las enzimas peroxisomales, el propionil-CoA (y el isobutiril-CoA) están implicados en el metabolismo de muchos sustratos diferentes (cuya identidad se está evaluando actualmente), y no sólo de la valina.

Metabolismo de los hongosEditar
La producción de propionil-CoA a través del catabolismo de los ácidos grasos también está asociada a la tioesterifcación. En un estudio relativo a Aspergillus nidulans, se comprobó que con la inhibición de un gen de la metilcitrato sintasa, mcsA, de la vía descrita anteriormente, se inhibía también la producción de distintos policétidos. Por lo tanto, la utilización de propionil-CoA a través del ciclo del metilcitrato disminuye su concentración, mientras que posteriormente aumenta la concentración de policétidos. Un policétido es una estructura comúnmente encontrada en los hongos que se compone de acetil-CoAs y malonil-CoAs, proporcionando un producto con grupos carbonilo y grupos metileno alternados. Los policétidos y sus derivados suelen ser muy complejos estructuralmente y algunos son muy tóxicos. Esto ha llevado a la investigación sobre la limitación de la toxicidad de los policétidos para los cultivos en la agricultura a través de los hongos fitopatógenos.
Propionilación de proteínasEditar
El propionil-CoA es también un sustrato para la modificación postraduccional de las proteínas al reaccionar con los residuos de lisina en las proteínas, una reacción llamada propionilación de proteínas. Debido a las similitudes estructurales del acetil-CoA y del propionil-CoA, se cree que la reacción de propionilación utiliza muchas de las mismas enzimas utilizadas para la acetilación de proteínas. Aunque las consecuencias funcionales de la propionilación de proteínas y actualmente no se entienden completamente, la propionilación in vitro de la enzima Propionil-CoA Sintetasa controla su actividad.