Introduction
Olen ollut onnekas saadessani osallistua kahteen suureen fysiologiseen tutkimusretkeen äärimmäisiin korkeuksiin. Ensimmäinen oli Silver Hut -retkikunta vuosina 1960-1961, jonka aikana ryhmä fysiologeja vietti useita kuukausia erittäin korkealla 5800 metrin korkeudessa, ja me mittasimme fysiologisia muutoksia, joita tapahtui tämän pitkän ajanjakson aikana. Lisätutkimuksia tehtiin aina 7 830 metriin (25 700 jalkaan) asti. Yleisenä tarkoituksena oli selvittää mekanismeja, joiden avulla ihmiset, jotka normaalisti elävät lähellä merenpinnan tasoa, reagoivat vakavaan hypoksiaan pitkän ajanjakson ajan.
Toinen retkikunta oli amerikkalainen lääketieteellinen tutkimusretkikunta Everestille, joka tapahtui vuonna 1981. Tässä fysiologinen tavoite oli hyvin erilainen. Tavoitteena oli saada ensimmäiset ihmisen fysiologiset tiedot Mount Everestin huipulla (8 848 m, 29 028 ft) sen selvittämiseksi, miten ihmiset, jotka normaalisti elävät merenpinnan tasolla, voivat selviytyä maailman korkeimman kohdan äärimmäisestä hypoksiasta.
Silver Hut Expedition
Tämän retkikunnan suunnitteli Sir Edmund Hillary, joka yhdessä Tenzing Norgayn kanssa saavutti ensimmäisenä ihmisenä Mount Everestin huipun seitsemän vuotta aiemmin. Hillary oli tehnyt yhteistyötä Griffith Pugh’n, korkean paikan fysiologin, kanssa tässä Everestin ensikiipeämisessä, ja molemmat miehet olivat erittäin kiinnostuneita akklimatisaatioprosessista, joka mahdollistaa sen, että ihmiset voivat kiivetä lähes merenpinnan tasolta hyvin korkeille paikoille.
Erityisen kiinnostukseni kohteena oli keuhkojen diffuusiokapasiteetti. Barcroft (1925) oli esittänyt, että liikunta suuressa korkeudessa johtaisi valtimoiden hypoksemiaan, koska diffuusio rajoittuu verikaasuesteen läpi. Tämän testaamiseksi järjestimme, että retkikunnan jäsenet työskentelivät maksimikapasiteetillaan polkupyöräergometrillä, ja mittasimme valtimoiden happisaturaation äskettäin saatavilla olleella korvaoksimetrillä (West ym., 1962). Havaitsimme, että valtimoveren happisaturaatio itse asiassa laski selvästi huolimatta alveolaarisen PO2:n noususta, kun työtehoa nostettiin. Tämä oli vahva todiste diffuusion rajoittumisesta näissä vakavan hypoksian olosuhteissa. Itselläni tehdyt kaksoismittaukset osoittivat, että happisaturaatio oli vain 33 % maksimaalisessa rasituksessa, mikä kuvasti hyvin vakavaa hypoksemiaa.
Määritimme myös hiilimonoksidin diffuusiokapasiteetin retken aikana ja osoitimme, että se tuskin muuttui (West, 1962). Pieni nousu saattoi selittyä kehittyneellä polysytemialla. Johtopäätöksenä oli, että verikaasuesteen ominaisuudet eivät muuttuneet pitkäaikaisessa altistumisessa vaikealle hypoksialle. Tämä oli ensimmäinen selkeä osoitus keuhkojen hapensiirron diffuusiorajoituksesta vaikean rasituksen aikana hyvin korkealla.
Myöhemmin retkikunnan aikana tehtiin mittauksia maksimaalisesta hapenottokyvystä rasituksen aikana äärimmäisen korkealla 7 440 metrin (24 400 jalan) korkeudessa (Pugh ym., 1964). Näiden tietojen ekstrapolointi Everestin huipun korkeuteen osoitti, että huipulle olisi mahdotonta päästä ilman lisähappea. Veritutkimukset osoittivat merkittävää polysytemiaa 5 800 metrin korkeudessa elävillä koehenkilöillä. Keskimääräinen hemoglobiinipitoisuus oli 19,6 g/dl ja hematokriitti 55,8 %. Oli näyttöä siitä, että hematokriitin alkuvaiheen nousu johtui pääasiassa plasmatilavuuden vähenemisestä, mutta myöhemmin punasolujen massa lisääntyi huomattavasti. 5800 metrin korkeudessa elävien henkilöiden elektrokardiogrammissa näkyi merkittävää oikean kammion hypertrofiaa, ja joissakin kuvauksissa rintakehän johtimissa esiintyi T-aaltojen inversiota, mikä oletettavasti osoitti sydänlihaksen vakavaa hypoksiaa (Milledge, 1963). Neuropsykometrisiä toimintoja mitattiin korttien lajittelun avulla, ja havaittiin, että lajittelunopeus väheni, mutta keskittymisen lisääntyessä koehenkilöt pystyivät lajittelemaan virheettömästi. Kaikki retkikunnan jäsenet laihtuivat vakavasti ja hellittämättä asuessaan 5800 metrin korkeudessa, ja laihtuminen oli 0,5-1,5 kiloa viikossa. Yleinen käsitys oli, että ihmisten ei olisi mahdollista elää loputtomiin näin korkealla (Pugh, 1962).
Amerikkalainen lääketieteellinen tutkimusretkikunta Everestille
Kuten edellä todettiin, tämän retkikunnan tavoitteena oli selvittää, miten ihminen voi sietää maailman korkeimman korkeuden hypoksiaa. Huomionarvoista oli, että muutamaa kuukautta ennen retkikuntaa kaksi eurooppalaista kiipeilijää saavutti Mount Everestin huipun ensimmäistä kertaa ilman lisähappea. Tämä saavutus hämmästytti monia fysiologeja ja herätti monia kysymyksiä siitä, miten se oli mahdollista.
Tutkimusohjelma oli hyvin laaja, ja tässä voidaan esittää vain lyhyt yhteenveto. Mittauksia tehtiin perusleirissä, korkeus 5400 m (17 700 ft), ja edistyneessä perusleirissä, korkeus 6300 m (20 700 ft), sekä korkeimmassa leirissä 8050 m (26 400 ft). Toivoimme myös saavamme mittauksia Everestin huipulla, vaikka tämä olikin hyvin kunnianhimoinen tavoite. Kun muistelimme kuutta omaa retkikuntaamme edeltänyttä retkikuntaa, yksikään niistä ei ollut päässyt huipulle. Jos sää on huono, unohda se, ja kriittinen tekijä on se, pysyvätkö riittävästi retkikunnan jäseniä riittävän hyväkuntoisina äärimmäisestä korkeudesta huolimatta.
Tukileirissä mittasimme hengitysreaktiota hypoksiaan eli sitä, missä määrin hengitys lisääntyy, kun koehenkilö altistuu vähäiselle hengitettävälle happiseokselle. Tulokset olivat hätkähdyttäviä. Kävi ilmi, että kiipeilijällä, joka pääsi huipulle ensimmäisenä, oli korkein vaste, toiseksi korkein vaste oli toiseksi korkeimmalla, ja kolmanneksi korkein vaste oli kiipeilijällä, joka pääsi huipulle kolmantena (Schoene ym., 1984). Tämän täytyy olla osittain sattumaa, mutta se viittaa varmasti siihen, että kiipeilijöiden ilmanvaihdon lisäämisen ja äärimmäisten korkeuserojen sietokyvyn välillä on yhteys. Syy tähän selviää tarkemmin jäljempänä.
Edistyneessä perusleirissä tehtiin lukuisia tutkimuksia, mutta tässä esitetään yhteenveto vain yhdestä, neuropsykometrisestä tutkimuksesta. On tunnettua, että aivot ja keskushermosto ovat hyvin herkkiä hypoksialle. Jos esimerkiksi joku putoaa uima-altaaseen ja hänet pelastetaan 5 tai 10 minuuttia myöhemmin, hänet saatetaan elvyttää onnistuneesti, mutta keskushermosto ei koskaan toivu täysin. Siksi ei ollut yllättävää, että pystyimme osoittamaan muutoksia mittauksissa, kuten lyhytkestoisessa muistissa ja manipulointitaidoissa (jotka määritettiin sormella naputtelukokeesta), hyvin suurissa korkeuksissa. Tämä ei ollut odottamatonta. Kun retkikunta kuitenkin palasi lähelle merenpinnan tasoa, havaittiin, että kaksi mittaustulosta oli edelleen epänormaaleja. Nämä olivat lyhytkestoinen muisti ja sorminäppäryystesti (Townes et al., 1984). Näin ollen oli selvää, että näiltä äärimmäisiltä korkeuksilta palaavilla henkilöillä on todennäköisesti jonkinlainen keskushermoston jäännöshäiriö. Olimme yksi ensimmäisistä ryhmistä, jotka osoittivat tämän, mutta se on sittemmin vahvistettu useaan otteeseen.
Joitakin mielenkiintoisimpia havaintoja saatiin huipulta. Olimme suunnitelleet ja rakentaneet erikoislaitteen, jonka avulla kiipeilijä pystyi keräämään maksimaalisen uloshengityksen jälkeen viimeisen uloshengityskaasun, eli alveolikaasunäytteen. Yli 34 näytettä, joista neljä huipulta, tuotiin takaisin UC San Diegoon kaasutiiviissä tölkeissä. Kun alveolaarinen PCO2 suhteutettiin barometriseen paineeseen, joka laski korkeuden kasvaessa, havaittiin, että PCO2 oli huipulla 7-8 mmHg. Tämä oli lähes uskomattoman alhainen arvo, koska merenpinnan taso on noin 40 mm Hg. Tämä äärimmäisen alhainen arvo korostaa alveolaarisen ventilaation valtavaa lisääntymistä, joka on välttämätöntä näissä äärimmäisissä korkeuksissa (West et al., 1983).
Kun sekä alveolaarinen PO2 että PCO2 piirrettiin barometristä painetta vastaan, saatiin mielenkiintoinen kuva (kuva 1). Sekä PO2 että PCO2 laskivat korkeuden kasvaessa. PO2:n lasku johtui kiipeilijää ympäröivän ilman vähenemisestä barometrisen paineen laskun seurauksena. PCO2:n lasku johtui ainoastaan kiipeilijän hyperventilaatiosta. Kävi ilmi, että kun korkeus ylitti noin 7000 metriä, alveolaarinen PO2 ei enää laskenut. Kuvasta näkyy, että tätä puolustettiin noin 35 mm Hg:n tasolla. Toisin sanoen näissä valtavissa korkeuksissa selviytyminen edellyttää alveolaarista ventilaatiota, joka laskee PCO2:n noin 8 mmHg:iin ja säilyttää näin alveolaarisen PO2:n hyvin alhaisella, mutta elinkelpoisella noin 35 mmHg:n tasolla. Tämä selittää sen, miksi edellä mainituissa hypoksiaan liittyvissä ventilaatiovasteen mittauksissa oli havaittavissa korrelaatio vasteen suuruuden ja kiipeilijän sietokyvyn välillä äärimmäisessä korkeudessa. Jos et kykene suorittamaan sellaista hyperventilaatiota, joka riittää alveolaarisen PCO2:n laskemiseen noin 7-8 mmHg:iin, et voi ylläpitää elinkelpoista PO2-tasoa alveolaarikaasussa. Näin ollen äärimmäinen hyperventilaatio on yksi tärkeimmistä piirteistä fysiologisessa vasteessa äärimmäisen korkealla.
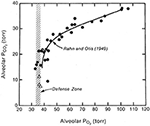
Kuva 1. Kiipeilijöiden alveolikaasun PO2- ja PCO2-arvot heidän noustessaan merenpinnan tasolta (ylhäällä oikealla) Everestin huipulle (alhaalla vasemmalla). PO2 laskee barometrisen paineen alenemisen vuoksi. PCO2 laskee, koska alveolaarinen ventilaatio lisääntyy. Kun saavutetaan korkeus, joka vastaa noin 20 mmHg:n PCO2:ta (noin 7000 m), PO2 ei enää laske. Toisin sanoen se puolustetaan noin 35 mmHg:n tasolla. Tämä onnistuu vain, jos PCO2:ta alennetaan jatkuvasti edelleen äärimmäisellä hyperventilaatiolla, kun korkeus nousee. Muokattu lähteistä Rahn ja Otis (1949) ja West ym. (1983).
Erittäin alhainen alveolaarinen PCO2 herättää kysymyksen siitä, mitä tapahtui valtimon pH:lle. On järkevää olettaa, että arteriaalinen ja alveolaarinen PCO2 ovat samat. Onneksi kaksi kiipeilijää otti toisiltaan laskimoverta onnistuneen huipulle kiipeämisen jälkeisenä aamuna, joten perusyliarvot voitiin mitata. Kun nämä arvot syötettiin Henderson-Hasselbalchin yhtälöön, tuloksena saatu valtimon pH oli 7,7-7,8. Tämä on äärimmäinen hengitysalkaloosi.
Mielenkiintoinen kysymys on, miksi munuaiset eivät poistaneet bikarbonaattia kehittääkseen metabolista kompensaatiota tälle äärimmäiselle alkaloosille ja siirtääkseen siten pH:ta lähemmäs normaalia. Tämä on tavanomainen reaktio, jos hengitysalkaloosi syntyy esimerkiksi hyperventilaation seurauksena, kuten joskus tapahtuu hysterian aikana. Syytä metabolisen kompensaation puuttumiseen ei täysin ymmärretä, mutta yksi mahdollisuus on se, että nämä kiipeilijät olivat luultavasti vakavasti tilavuuspuutteessa. Tämä on yleinen piirre, kun mennään suuriin korkeuksiin, ja esimerkiksi edistyneessä perusleirissä 6300 metrin korkeudessa olleilla kiipeilijöillä oli merkkejä kroonisesta tilavuusvajeesta. Yksi vastuullinen tekijä äärimmäisessä korkeudessa on oletettavasti valtava hyperventilaatio, mutta myös jano on heikentynyt.
Vaikean alkaloosin fysiologiset seuraukset ovat mielenkiintoisia. Toiset tutkimukset ovat osoittaneet, että hemoglobiinin lisääntynyt happiaffiniteetti lisää selviytymistä hypoksisessa ympäristössä. Monta vuotta sitten osoitettiin, että Etelä-Amerikan Andeilla elävillä nisäkkäillä, kuten vicuñalla ja laamalla, on kohonnut happiaffiniteetti (eli niillä on vasemmalle -siirtynyt hapen dissosiaatiokäyrä) verrattuna merenpinnan tasolla eläviin nisäkkäisiin (Hall, 1937). Näin ollen hyvin korkealla kiipeilijöillä on sama vaste.
On myös totta, että jos tarkastellaan yleisesti koko eläinkunnassa organismeja, jotka elävät hypoksisessa ympäristössä, monille niistä on kehittynyt lisääntynyt hemoglobiinin happiaffiniteetti. Yksi parhaista esimerkeistä on ihmisen sikiö, jonka hemoglobiinin aminohapposekvenssin eron vuoksi happiaffiniteetti on selvästi kohonnut P50:n ollessa noin 19 mmHg verrattuna aikuisen noin 27 mmHg:n P50:een. Ihmissikiöllä on aikuisen standardien mukaan vakava hypoksemia, sillä PO2 laskevassa aortassa on noin 22 mmHg, mikä on jopa alhaisempi kuin Everestin huipulla kiipeävällä kiipeilijällä. On todellakin kiehtovaa, että onnistuneella kiipeilijällä on se etu, että hemoglobiinin happiaffiniteetti on kasvanut. Tämä auttaa hapen latautumista keuhkokapillaariin. Voidaan väittää, että se haittaa myös hapen purkautumista kehon perifeerisissä kudoksissa, mutta tutkimukset ovat osoittaneet, että keuhkoissa tapahtuvan latautumisen etu on suurempi kuin perifeerisissä kudoksissa tapahtuvan purkautumisen haitta.
Mielenkiintoinen kysymys on, mikä on kiipeilijän maksimaalinen hapenkulutus huipulla. Kuten aiemmin todettiin, aiemmat mittaukset, jotka tehtiin äärimmäisessä korkeudessa Silver Hut -retkikunnan aikana, viittasivat siihen, että kaikki huipulla käytettävissä oleva happi tarvittaisiin perushapen päivitykseen, eli sydämen pumppaamiseen ja aivojen aktiivisena pitämiseen. Pyöräergometriä oli mahdotonta sijoittaa huipulle. Otimme kuitenkin kiipeilijöitä, jotka olivat hyvin akklimatisoituneet, ja panimme heidät polkemaan maksimaalisesti 6300 metrin korkeudessa hengittäen 14 prosenttia happea. Näin he saivat saman innostuneen PO2:n kuin huipulla. Hapenottotaso näissä olosuhteissa oli noin 1 l/min, mikä on surkean alhainen maksimaalinen hapenkulutus ja vastaa sitä, että joku kävelee hitaasti tasamaalla. Ilmeisesti se kuitenkin riittää juuri ja juuri siihen, että kiipeilijä pääsee huipulle.
Author Contributions
JW laati käsikirjoituksen, luki ja hyväksyi toimitetun version.
Conflict of Interest Statement
Tekijä vakuuttaa, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Barcroft, J. (1925). Veren hengitystoiminta. Osa 1 Lessons from High Altitudes. Cambridge: Cambridge University Press.
Google Scholar
Hall, F. G. (1937). Eläinten sopeutuminen suuriin korkeuksiin. J. Mammol. 18, 469-472. doi: 10.2307/1374337
CrossRef Full Text | Google Scholar
Milledge, J. S. (1963). Elektrokardiografiset muutokset suuressa korkeudessa. Br. Heart J. 15, 291-298. doi: 10.1136/hrt.25.3.291
CrossRef Full Text | Google Scholar
Pugh, L. G. (1962). Himalajan ja tieteellisen retkikunnan 1960-61 fysiologiset ja lääketieteelliset näkökohdat. Br. Med. J. 2, 621-633. doi: 10.1136/bmj.2.5305.621
CrossRef Full Text | Google Scholar
Pugh, L. G., Gill, M. B., Lahiri, S., Milledge, J. S., Ward, M. P., ja West, J. B. (1964). Lihaskuntoharjoittelu suurilla korkeuksilla. J. Appl. Physiol 19, 431-440. doi: 10.1152/jappl.1964.19.3.431
CrossRef Full Text | Google Scholar
Rahn, H., and Otis, A. B. (1949). Ihmisen hengitysvaste korkeuteen akklimatisoitumisen aikana ja sen jälkeen. Am. J. Physiol. 157, 445-462. doi: 10.1152/ajplegacy.1949.157.3.445
PubMed Abstract | CrossRef Full Text | Google Scholar
Schoene, R. B., Lahiri, S., Hackett, P. H., Peters, R. M., Milledge, J. S., Pizzo, C. J., et al. (1984). Hypoksisen ventilaatiovasteen suhde liikuntasuoritukseen Mount Everestillä. J. Appl. Physiol. 56, 1478-1483. doi: 10.1152/jappl.1984.56.6.1478
CrossRef Full Text | Google Scholar
Townes, B. D., Hornbein, T. H., and Grant, I. (1984). ”Human cerebral function at extreme altitude,” in High Altitude and Man, eds J. B. West and S. Lahiri (Bethesda, MD: American Physiological Society), 31-36.
Google Scholar
West, J. B. (1962). Keuhkojen diffuusiokapasiteetti hiilimonoksidille korkealla. J. Appl. Physiol. 17, 421-426. doi: 10.1152/jappl.1962.17.3.421
CrossRef Full Text | Google Scholar
West, J. B., Hackett, P. H., Maret, K. H., Milledge, J. S., Peters, R. M., Pizzo, C. J., et al. (1983). Keuhkojen kaasunvaihto Mount Everestin huipulla. J. Appl. Physiol. 55:678-687. doi: 10.1152/jappl.1983.55.3.678
CrossRef Full Text | Google Scholar
West, J. B., Lahiri, S., Gill, M. B., Milledge, J. S., Pugh, L. G. C. E., ja Ward, M. P. (1962). Arteriaalinen hapenotto liikunnan aikana suuressa korkeudessa. J. Appl. Physiol. 17, 617-621. doi: 10.1152/jappl.1962.17.4.617
CrossRef Full Text | Google Scholar