Wprowadzenie
Miałem szczęście uczestniczyć w dwóch dużych fizjologicznych wyprawach badawczych na ekstremalne wysokości. Pierwszą z nich była ekspedycja Silver Hut w latach 1960-1961, podczas której grupa fizjologów spędziła kilka miesięcy na bardzo dużej wysokości 5.800 m (19.000 stóp), a my mierzyliśmy zmiany fizjologiczne, które zachodziły w tym długim okresie. Dodatkowe badania zostały przeprowadzone do wysokości 7.830 m (25.700 stóp). Ogólnym celem było określenie mechanizmów, dzięki którym ludzie, którzy normalnie żyją w pobliżu poziomu morza, reagują na ciężką hipoksję przez dłuższy okres.
Drugą wyprawą była American Medical Research Expedition to Everest, która miała miejsce w 1981 roku. Tutaj cel fizjologiczny był zupełnie inny. Celem było uzyskanie pierwszych ludzkich danych fizjologicznych na szczycie Mount Everest (8,848 m, 29,028 ft) w celu wyjaśnienia, jak ludzie, którzy normalnie żyją na poziomie morza może przetrwać skrajne niedotlenienie najwyższego punktu na świecie.
Silver Hut Expedition
To był pomysł Sir Edmunda Hillary’ego, który wraz z Tenzing Norgay, był pierwszą osobą, która osiągnęła szczyt Mt Everest siedem lat wcześniej. Hillary współpracował z Griffith Pugh, fizjolog wysokiej wysokości, na tym pierwszym wejściu Everest, a obaj mężczyźni byli intensywnie zainteresowani w procesie aklimatyzacji, który umożliwia ludziom z blisko poziomu morza, aby wspiąć się na bardzo high altitudes.
Moje szczególne zainteresowanie było dyfuzji pojemność płuc. To zostało zasugerowane przez Barcroft (1925), że ćwiczenia na dużych wysokościach spowoduje hipoksemii tętniczej z powodu ograniczenia dyfuzji przez barierę krew-gaz. Aby to sprawdzić, zorganizowaliśmy dla członków ekspedycji pracę z maksymalną wydajnością na ergometrze rowerowym i zmierzyliśmy nasycenie tlenem tętniczym przy użyciu nowo dostępnego oksymetru dousznego (West et al., 1962). Stwierdziliśmy, że w rzeczywistości nastąpił wyraźny spadek saturacji tlenem tętniczym, pomimo wzrostu PO2 w pęcherzykach płucnych, gdy poziom pracy został zwiększony. Był to mocny dowód na ograniczenie dyfuzji w tych warunkach ciężkiej hipoksji. Powielone pomiary na sobie wykazały nasycenie tlenem tylko 33% przy maksymalnym wysiłku, co odzwierciedlało bardzo ciężką hipoksemię.
Pomierzyliśmy również zdolność dyfuzyjną dla tlenku węgla w trakcie wyprawy i wykazaliśmy, że prawie się nie zmieniła (West, 1962). Niewielki wzrost może być wyjaśnione przez polythemia, która rozwinęła. Wniosek był taki, że cechy bariery gazowej krwi nie zostały zmienione przez długotrwałe narażenie na ciężką hipoksję. Było to pierwsze wyraźne wykazanie dyfuzji ograniczenie transferu tlenu przez płuco podczas ciężkich ćwiczeń na bardzo dużej wysokości.
Później w ekspedycji, pomiary zostały wykonane z maksymalnego poboru tlenu podczas ćwiczeń na bardzo dużej wysokości 7,440 m (24,400 ft) (Pugh et al., 1964). Ekstrapolacja tych danych do wysokości szczytu Everest sugeruje, że byłoby niemożliwe, aby osiągnąć szczyt bez dodatkowego tlenu. Badania krwi wykazały wyraźną policytemię u osób żyjących na wysokości 5800 m. Średnie stężenie hemoglobiny i hematokryt wynosiły odpowiednio 19,6 g/dl i 55,8%. Istniały dowody, że początkowy wzrost hematokrytu był spowodowany głównie utratą objętości osocza, ale później nastąpił duży wzrost masy krwinek czerwonych. Elektrokardiogram osób żyjących na wysokości 5800 m n.p.m. wykazywał wyraźny przerost prawej komory, a w niektórych zapisach występowała inwersja załamka T w odprowadzeniach piersiowych, co prawdopodobnie wskazywało na poważne niedotlenienie mięśnia sercowego (Milledge, 1963). Pomiary funkcji neuropsychometrycznych przeprowadzono za pomocą sortowania kart i stwierdzono, że szybkość sortowania była zmniejszona, ale przy zwiększonej koncentracji badani mogli sortować bezbłędnie. Nie było poważne, nieubłagany spadek masy ciała we wszystkich członków ekspedycji podczas życia na 5.800 m z szybkością między 0,5 i 1,5 Kg / tydzień. Ogólne wrażenie było, że nie byłoby możliwe dla ludzi, aby żyć w nieskończoność na tej bardzo dużej wysokości (Pugh, 1962).
American Medical Research Expedition to Everest
Jak wspomniano powyżej, celem tej ekspedycji było wyjaśnienie, jak ludzie mogą tolerować hipoksję najwyższej wysokości na świecie. Co niezwykłe, kilka miesięcy przed wyprawą, dwóch europejskich wspinaczy osiągnęło szczyt Mount Everest po raz pierwszy bez użycia dodatkowego tlenu. Ten wyczyn zadziwił wielu fizjologów, i podniósł wiele pytań o to, jak to może być done.
Program badawczy był bardzo obszerny, a tylko krótkie streszczenie może być podane tutaj. Pomiary zostały wykonane w obozie bazowym, wysokość 5,400 m (17,700 ft), i zaawansowanym obozie bazowym, wysokość 6,300 m (20,700 ft), i w najwyższym obozie 8,050 m (26,400 ft). Mieliśmy również nadzieję, że uda nam się wykonać kilka pomiarów na szczycie Everestu, choć było to bardzo ambitne zadanie. W rzeczywistości, gdy spojrzeliśmy wstecz na sześć ekspedycji poprzedzających naszą, żadna z nich nie zdobyła szczytu. Jeśli pogoda jest zła, zapomnij o tym, a krytycznym czynnikiem jest to, czy wystarczająca liczba członków ekspedycji pozostaje wystarczająco sprawny pomimo ekstremalnej wysokości.
W obozie bazowym, zmierzyliśmy odpowiedź wentylacyjną na hipoksję, czyli zakres, w jakim oddychanie wzrasta, gdy podmiot jest narażony na niską natchnionej mieszaniny tlenu. Wyniki były uderzające. Okazało się, że wspinacz, który dotarł na szczyt jako pierwszy miał najwyższą odpowiedź, ten, który dotarł na szczyt jako drugi miał kolejną najwyższą odpowiedź, a wspinacz, który dotarł na szczyt jako trzeci miał trzecią najwyższą odpowiedź (Schoene i in., 1984). To musi być częściowo przez przypadek, ale na pewno sugeruje, że istniał związek między stopniem, do którego wspinacze zwiększyć ich wentylacji, a ich tolerancji na ekstremalnych wysokościach. Przyczyna tego stanie się jaśniejsza poniżej.
Duża liczba badań została przeprowadzona w zaawansowanym obozie bazowym, ale tylko jedno, badanie neuropsychometryczne, zostanie podsumowane tutaj. Powszechnie wiadomo, że mózg i centralny układ nerwowy są bardzo wrażliwe na niedotlenienie. Na przykład, jeśli ktoś wpadnie do basenu i zostanie uratowany 5 lub 10 minut później, może być skutecznie ożywiony, ale centralny układ nerwowy nigdy nie odzyskuje pełnej sprawności. Nie było więc zaskoczeniem, że mogliśmy wykazać zmiany w pomiarach takich jak pamięć krótkotrwała i umiejętności manipulacyjne (określone na podstawie testu stukania palcami) na bardzo dużych wysokościach. Nie było to niespodziewane. Jednakże, kiedy ekspedycja powróciła na poziom zbliżony do poziomu morza, okazało się, że dwa z pomiarów pozostały nienormalne. Były to pamięć krótkotrwała i test stukania palcami (Townes et al., 1984). Było więc jasne, że każda osoba powracająca z tych ekstremalnych wysokości prawdopodobnie będzie miała jakieś resztkowe upośledzenie centralnego układu nerwowego. Byliśmy jedną z pierwszych grup, aby pokazać to, ale to zostało potwierdzone wiele razy od.
Kilka z najbardziej interesujących odkryć były te z szczytu. Zaprojektowaliśmy i zbudowaliśmy specjalne urządzenie, które umożliwiło wspinaczowi zebranie ostatniego wydychanego gazu po maksymalnym wydechu, czyli próbki gazu pęcherzykowego. Ponad 34 próbki, w tym cztery ze szczytu, zostały przywiezione do UC San Diego w gazoszczelnych puszkach. Kiedy PCO2 z pęcherzyków płucnych zostało porównane z ciśnieniem barometrycznym, które spadało wraz ze wzrostem wysokości, okazało się, że PCO2 na szczycie wynosiło 7-8 mm Hg. Była to niemal niewiarygodnie niska wartość, ponieważ wartość na poziomie morza wynosi około 40 mm Hg. Ta niezwykle niska wartość podkreśla ogromny wzrost wentylacji pęcherzykowej, która jest niezbędna na tych ekstremalnych wysokościach (West et al., 1983).
Gdy zarówno pęcherzykowe PO2 i PCO2 zostały wykreślone w stosunku do ciśnienia barometrycznego, i ciekawy obraz wyłonił się (rysunek 1). Zarówno PO2 jak i PCO2 spadały wraz ze wzrostem wysokości nad poziomem morza. Spadek PO2 był spowodowany zmniejszeniem ilości powietrza wokół wspinacza w wyniku obniżenia ciśnienia barometrycznego. Spadek PCO2 był spowodowany wyłącznie hiperwentylacją wspinacza. Okazało się, że po przekroczeniu wysokości około 7000 m n.p.m. nie dochodzi do dalszego spadku pęcherzykowego PO2. Rysunek pokazuje, że jest to bronione na poziomie około 35 mm Hg. Innymi słowy, aby przetrwać na tych ogromnych wysokościach, trzeba zamontować wentylację pęcherzyków płucnych, która obniży PCO2 do około 8 mmHg i w ten sposób utrzyma PO2 pęcherzyków płucnych na bardzo niskim, ale możliwym do utrzymania poziomie około 35 mm Hg. To wyjaśnia, dlaczego w pomiarach odpowiedzi oddechowej na hipoksję, o których była mowa wcześniej, istniała korelacja pomiędzy wielkością odpowiedzi a tolerancją wspinacza na ekstremalną wysokość. Jeśli nie jesteś w stanie zamontować takiego stopnia hiperwentylacji, który jest wystarczający do obniżenia PCO2 w pęcherzykach płucnych do około 7-8 mm Hg, nie możesz utrzymać realnego poziomu PO2 w gazie pęcherzykowym. Tak więc, skrajna hiperwentylacja jest jedną z najważniejszych cech fizjologicznej odpowiedzi na ekstremalnie dużą wysokość.
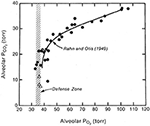
Rysunek 1. Wartości PO2 i PCO2 w gazie pęcherzykowym alpinistów w trakcie wznoszenia się z poziomu morza (u góry po prawej) na szczyt Everestu (u dołu po lewej). PO2 spada z powodu obniżenia ciśnienia barometrycznego. PCO2 spada z powodu zwiększonej wentylacji pęcherzyków płucnych. Powyżej wysokości odpowiadającej PCO2 około 20 mm Hg (około 7000 m), nie ma dalszego spadku PO2. Innymi słowy, jest ono bronione na poziomie około 35 mmHg. Może to nastąpić tylko wtedy, gdy PCO2 jest stale obniżane przez ekstremalną hiperwentylację w miarę wzrostu wysokości. Zmodyfikowane z Rahn i Otis (1949) i West et al. (1983).
Niezwykle niskie pęcherzykowe PCO2 skłania do pytania, co stało się z tętniczym pH. Rozsądnie jest założyć, że tętnicze i pęcherzykowe PCO2 są takie same. Na szczęście dwóch wspinaczy pobrało sobie nawzajem krew żylną rano po udanym wejściu na szczyt, dzięki czemu można było zmierzyć podstawowe wartości nadmiaru. Kiedy wartości te zostały wprowadzone do równania Hendersona-Hasselbalcha, otrzymane pH tętnicze wynosiło pomiędzy 7,7 a 7,8. Jest to ekstremalny stopień alkalozy oddechowej.
Ciekawe pytanie brzmi, dlaczego nerki nie wyeliminowały wodorowęglanu, aby rozwinąć metaboliczną kompensację dla tej ekstremalnej alkalozy, a tym samym przesunąć pH bliżej normy. Jest to zwykła odpowiedź, jeśli alkaloza oddechowa jest generowana, na przykład, przez hiperwentylację, jak czasami występuje podczas histerii. Przyczyna braku kompensacji metabolicznej nie jest całkowicie zrozumiała, ale możliwe jest, że ci wspinacze byli prawdopodobnie poważnie pozbawieni objętości. Jest to powszechna cecha pójścia na dużą wysokość i, na przykład, wspinacze, którzy przebywali w zaawansowanym obozie bazowym na 6300 m wykazały dowody przewlekłego wyczerpania objętości. Jednym z odpowiedzialnych czynników na ekstremalnych wysokościach jest przypuszczalnie ogromny hiperwentylacji, ale pragnienie jest upośledzone, jak również.
Fizjologiczne konsekwencje ciężkiej alkalozy są interesujące. Inne badania wykazały, że zwiększone powinowactwo tlenu hemoglobiny zwiększa przeżycie w hipoksyjnym środowisku. Wiele lat temu wykazano, że ssaki takie jak vicuña i lama w południowoamerykańskich Andach mają zwiększone powinowactwo do tlenu (czyli miały przesuniętą w lewo krzywą dysocjacji tlenu) w porównaniu ze ssakami żyjącymi na poziomie morza (Hall, 1937). Tak więc, wspinaczy na bardzo dużej wysokości mają taką samą odpowiedź.
Jest również prawdą, że jeśli spojrzeć ogólnie w całym królestwie zwierząt na organizmy, które żyją w środowisku hipoksycznym, wiele z nich rozwinęło zwiększone powinowactwo tlenu hemoglobiny. Jednym z najlepszych przykładów jest płód ludzki, który, z powodu różnicy w sekwencji aminokwasów hemoglobiny, ma wyraźny wzrost powinowactwa tlenu z P50 około 19 mm Hg w porównaniu z tym około 27 dla dorosłego. Płód ludzki ma ciężką hipoksemię według standardów dorosłych z PO2 w aorcie zstępującej około 22 mm Hg, który jest nawet niższy niż wspinacza na szczycie Everestu. Fascynujące jest to, że udany wspinacz ma przewagę zwiększonego powinowactwa hemoglobiny do tlenu. Pomaga to w ładowaniu tlenu w kapilarach płucnych. Można argumentować, że to również koliduje z rozładowania tlenu w peryferiach ciała, ale badania wykazały, że korzyść z załadunku w płucach jest większa niż wada w rozładunku w tkankach peryferyjnych.
Ciekawe pytanie jest to, co jest maksymalne zużycie tlenu przez wspinacza na szczycie. Jak wskazano wcześniej, poprzednie pomiary wykonane na ekstremalnych wysokościach podczas ekspedycji Srebrna Chata sugerowały, że cały tlen dostępny na szczycie byłby wymagany do podstawowej aktualizacji tlenu, to jest do utrzymania serca pompowania i mózg aktywny. Umieszczenie ergometru rowerowego na szczycie było niemożliwe. Wzięliśmy jednak wspinaczy, którzy byli bardzo dobrze zaaklimatyzowani i kazaliśmy im pedałować maksymalnie na wysokości 6300 m, oddychając 14% tlenem. To dało im takie samo natchnione PO2 jak na szczycie. Poziom poboru tlenu w tych warunkach wynosił około 1 L/min, co jest żałośnie niskim maksymalnym zużyciem tlenu i odpowiada powolnemu spacerowi na poziomie. Jest to jednak najwyraźniej wystarczające, aby umożliwić wspinaczowi zdobycie szczytu.
Wkład autorów
JW sporządził manuskrypt, przeczytał i zatwierdził przedłożoną wersję.
Conflict of Interest Statement
Autor oświadcza, że badania zostały przeprowadzone przy braku jakichkolwiek komercyjnych lub finansowych powiązań, które mogłyby być interpretowane jako potencjalny konflikt interesów.
Barcroft, J. (1925). The Respiratory Function of the Blood. Part 1 Lessons from High Altitudes. Cambridge: Cambridge University Press.
Google Scholar
Hall, F. G. (1937). Adaptacje zwierząt do przebywania na dużych wysokościach. J. Mammol. 18, 469-472. doi: 10.2307/1374337
CrossRef Full Text | Google Scholar
Milledge, J. S. (1963). Zmiany elektrokardiograficzne na dużych wysokościach. Br. Heart J. 15, 291-298. doi: 10.1136/hrt.25.3.291
CrossRef Full Text | Google Scholar
Pugh, L. G. (1962). Fizjologiczne i medyczne aspekty ekspedycji himalajskiej i naukowej 1960-61. Br. Med. J. 2, 621-633. doi: 10.1136/bmj.2.5305.621
CrossRef Full Text | Google Scholar
Pugh, L. G., Gill, M. B., Lahiri, S., Milledge, J. S., Ward, M. P., and West, J. B. (1964). Muscular exercise at great altitudes. J. Appl. Physiol 19, 431-440. doi: 10.1152/jappl.1964.19.3.431
CrossRef Full Text | Google Scholar
Rahn, H., and Otis, A. B. (1949). Man’s respiratory response during and after acclimatization to high altitude. Am. J. Physiol. 157, 445-462. doi: 10.1152/ajplegacy.1949.157.3.445
PubMed Abstract | CrossRef Full Text | Google Scholar
Schoene, R. B., Lahiri, S., Hackett, P. H., Peters, R. M., Milledge, J. S., Pizzo, C. J., et al. (1984). Relationship of hypoxic ventilatory response to exercise performance on Mt Everest. J. Appl. Physiol. 56, 1478-1483. doi: 10.1152/jappl.1984.56.6.1478
CrossRef Full Text | Google Scholar
Townes, B. D., Hornbein, T. H., and Grant, I. (1984). „Human cerebral function at extreme altitude,” in High Altitude and Man, eds J. B. West and S. Lahiri (Bethesda, MD: American Physiological Society), 31-36.
Google Scholar
West, J. B. (1962). Pojemność dyfuzyjna płuc dla tlenku węgla na dużych wysokościach. J. Appl. Physiol. 17, 421-426. doi: 10.1152/jappl.1962.17.3.421
CrossRef Full Text | Google Scholar
West, J. B., Hackett, P. H., Maret, K. H., Milledge, J. S., Peters, R. M., Pizzo, C. J., et al. (1983). Pulmonary gas exchange on the summit of Mt Everest. J. Appl. Physiol. 55:678-687. doi: 10.1152/jappl.1983.55.3.678
CrossRef Full Text | Google Scholar
West, J. B., Lahiri, S., Gill, M. B., Milledge, J. S., Pugh, L. G. C. E., and Ward, M. P. (1962). Arterial oxygenation during exercise at high altitude. J. Appl. Physiol. 17, 617-621. doi: 10.1152/jappl.1962.17.4.617
CrossRef Full Text | Google Scholar
.