Teren metaboliczny (kataboliczny) propionyl-CoA zależy od środowiska, w którym jest syntetyzowany. Dlatego propionyl-CoA w środowisku beztlenowym może mieć inny los niż w organizmie tlenowym. Wiele ścieżek, albo katabolizm przez karboksylazę propionyl-CoA lub syntazę metylocitratu, również zależy od obecności różnych genów.
Reakcja z karboksylazą propionyl-CoAEdit
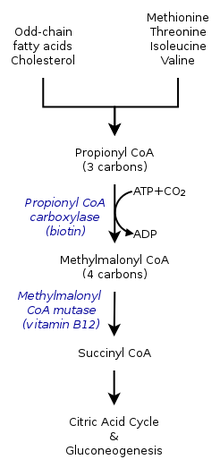
W cyklu kwasu cytrynowego u ludzi, propionyl-CoA, który oddziałuje z oksalooctanem w celu utworzenia metylocitratu, może być również katalizowany do metylomalonylo-CoA poprzez karboksylację przez karboksylazę propionyl-CoA (PCC). Metylomalonylo-CoA jest następnie przekształcany do sukcynylo-CoA, który jest dalej wykorzystywany w cyklu kwasu trójkarboksylowego. PCC nie tylko katalizuje karboksylację propionylo-CoA do metylomalonylo-CoA, ale także oddziałuje na kilka różnych acylo-CoA. Niemniej jednak, jego najwyższe powinowactwo wiążące jest do propionyl-CoA. Wykazano ponadto, że przemiana propionylo-CoA jest hamowana przez brak kilku markerów TCA, takich jak glutaminian. Mechanizm przedstawiono na rysunku po lewej stronie.
MechanismEdit
W ssakach, propionylo-CoA jest przekształcany do (S)-metylomalonylo-CoA przez karboksylazę propionylo-CoA, enzym zależny od biotyny, wymagający również wodorowęglanu i ATP.
Ten produkt jest przekształcany do (R)-metylomalonylo-CoA przez racemazę metylomalonylo-CoA.
(R)-metylomalonylo-CoA jest przekształcany do sukcynylo-CoA, produktu pośredniego w cyklu kwasu trójkarboksylowego, przez mutazę metylomalonylo-CoA, enzym wymagający

kobalaminy do katalizowania migracji wiązania węgiel-węgiel.
Mechanizm mutazy metylomalonylo-CoA rozpoczyna się od rozszczepienia wiązania między 5′ CH
2- z 5′-deoksyadenozylu a kobaltem, który znajduje się w stanie utlenienia 3+ (III), co powoduje powstanie rodnika 5′-deoksyadenozylu i kobalaminy w zredukowanym stanie utlenienia Co(II).
Następnie, rodnik ten abstrahuje atom wodoru z grupy metylowej metylomalonylo-CoA, co generuje rodnik metylomalonylo-CoA. Uważa się, że ten rodnik tworzy wiązanie węgiel-kobalt do koenzymu, który jest następnie przez rearanżację szkieletu węglowego substratu, wytwarzając w ten sposób rodnik sukcynylo-CoA. Rodnik ten następnie przechodzi do abstrakcji wodoru z wcześniej wytworzonej 5′-deoksyadenozyny, ponownie tworząc rodnik deoksyadenozylowy, który atakuje koenzym, aby zreformować początkowy kompleks.
Wada enzymu mutazy metylomalonylo-CoA powoduje kwasicę metylomalonową, niebezpieczne zaburzenie, które powoduje obniżenie pH krwi.

Cykl metylocytrynowyEdit
Kumulacja propionyl-CoA może okazać się toksyczna dla różnych organizmów. Ponieważ zaproponowano różne cykle dotyczące tego, jak propionyl-CoA jest przekształcany w pirogronian, jednym z badanych mechanizmów jest cykl metylocytrynowy.początkową reakcją jest beta-oksydacja w celu utworzenia propionyl-CoA, który jest dalej rozkładany przez cykl. Ta ścieżka obejmuje enzymy zarówno związane z cyklem metylocytrynianowym, jak i cyklem kwasu cytrynowego. Wszystkie one przyczyniają się do ogólnej reakcji detoksykacji bakterii ze szkodliwego propionyl-CoA. Jest on również przypisywany jako ścieżka wynikowa w związku z katabolizmem kwasów tłuszczowych w prątkach. W celu kontynuacji, gen prpC koduje syntazę metylocitratu, a jeśli nie jest obecny, cykl metylocitratu nie wystąpi. Zamiast tego, katabolizm zachodzi poprzez karboksylazę propionyl-CoA. Mechanizm ten jest przedstawiony poniżej po lewej stronie wraz z uczestniczącymi reaktorami, produktami, produktami pośrednimi i enzymami.
Metabolizm bakteryjnyEdit
Metabolizm Mycobacterium tuberculosisEdit
Utlenianie propionylo-CoA do postaci pirogronianu jest uwarunkowane jego koniecznością w Mycobacterium tuberculosis. Akumulacja propionylo-CoA może prowadzić do efektów toksycznych. W Mycobacterium tuberculosis sugeruje się, że metabolizm propionylo-CoA jest zaangażowany w biogenezę ściany komórkowej. Brak takiego katabolizmu zwiększałby zatem podatność komórki na działanie różnych toksyn, szczególnie na mechanizmy przeciwdrobnoustrojowe makrofagów. Inna hipoteza dotycząca losu propionyl-CoA, w M. tuberculosisis, jest taka, że ponieważ propionyl-CoA jest produkowany przez katabolizm kwasów tłuszczowych o łańcuchu beta nieparzystym, cykl metylocytrynianowy jest następnie aktywowany w celu zanegowania jakiejkolwiek potencjalnej toksyczności, działając jako mechanizm buforujący.
Możliwa sekwestracja w R. sphaeroidesEdit
Propionyl-CoA może mieć wiele niekorzystnych i toksycznych wpływów na różne gatunki, w tym bakterie. Na przykład, hamowanie dehydrogenazy pirogronianowej przez nagromadzenie propionyl-CoA w Rhodobacter sphaeroides może okazać się śmiertelne. Ponadto, podobnie jak w przypadku E. coli, napływ propionylo-CoA w gatunkach Myobacterial może spowodować toksyczność, jeśli nie zajmiemy się tym natychmiast. Toksyczność ta jest spowodowana przez szlak obejmujący lipidy, które tworzą ścianę komórkową bakterii. Poprzez estryfikację długołańcuchowych kwasów tłuszczowych, nadmiar propionyl-CoA może być sekwestrowany i przechowywany w lipidach triacylogliceroli (TAG), co prowadzi do regulacji podwyższonego poziomu propionyl-CoA. Taki proces rozgałęziania metylowego kwasów tłuszczowych powoduje, że działają one jako pochłaniacze dla gromadzącego się propionu
Escherichia coli metabolismEdit
W badaniu przeprowadzonym przez Luo i wsp. szczepy Escherichia coli zostały wykorzystane do zbadania, jak metabolizm propionylo-CoA może potencjalnie prowadzić do produkcji kwasu 3-hydroksypropionowego (3-HP). Wykazano, że mutacja w kluczowym genie zaangażowanym w ten szlak, transferze bursztynianowej CoA, prowadziła do znacznego wzrostu 3-HP. Jest to jednak wciąż rozwijająca się dziedzina i informacje na ten temat są ograniczone.

Metabolizm roślinnyEdit
Metabolizm aminokwasów w roślinach został uznany za kontrowersyjny temat, ze względu na brak konkretnych dowodów na istnienie jakiejkolwiek konkretnej ścieżki. Jednakże sugeruje się, że enzymy związane z produkcją i wykorzystaniem propionyl-CoA są zaangażowane. Związany z tym jest metabolizm izobutyrylo-CoA. Te dwie cząsteczki są uważane za produkty pośrednie w metabolizmie waliny. Ponieważ propionian występuje w formie propionyl-CoA, odkryto, że propionyl-CoA jest przekształcany do β-hydroksypropionianu poprzez peroksysomalną enzymatyczną ścieżkę β-oksydacji. Jednakże u rośliny Arabidopsis nie zaobserwowano kluczowych enzymów w konwersji waliny do propionyl-CoA. Poprzez różne eksperymenty przeprowadzone przez Lucasa i wsp. zasugerowano, że w roślinach, poprzez enzymy peroksysomalne, propionylo-CoA (i izobutyrylo-CoA) są zaangażowane w metabolizm wielu różnych substratów (obecnie ocenianych pod kątem tożsamości), a nie tylko waliny.

Metabolizm grzybówEdit
Produkcja propionylo-CoA poprzez katabolizm kwasów tłuszczowych jest również związana z tioestryfikacją. W badaniu dotyczącym Aspergillus nidulans stwierdzono, że przy zahamowaniu genu syntazy metylocitratu, mcsA, opisanego powyżej szlaku, zahamowana została również produkcja wyraźnych poliketydów. Zatem utylizacja propionyl-CoA przez cykl metylocytrynianowy powoduje zmniejszenie jego stężenia, a następnie zwiększenie stężenia poliketydów. Poliketyd jest strukturą powszechnie występującą u grzybów, która zbudowana jest z acetylo- i malonylo-CoA, dając produkt z naprzemiennie występującymi grupami karbonylowymi i metylenowymi. Poliketidy i pochodne poliketydów są często bardzo złożone strukturalnie, a niektóre z nich są bardzo toksyczne. Doprowadziło to do badań nad ograniczeniem toksyczności polyketydów dla upraw w rolnictwie poprzez grzyby fitopatogenne.
Propionylacja białekEdit
Propionyl-CoA jest również substratem do potranslacyjnej modyfikacji białek poprzez reakcję z resztami lizyny na białkach, reakcję zwaną propionylacją białek. Ze względu na strukturalne podobieństwo Acetylo-CoA i Propionylo-CoA, uważa się, że w reakcji propionylacji białek bierze udział wiele z tych samych enzymów, które są wykorzystywane do acetylacji białek. Chociaż funkcjonalne konsekwencje propionylacji białek nie są obecnie w pełni poznane, to jednak w warunkach in vitro propionylacja enzynomu syntetazy Propionyl-CoA kontroluje jego aktywność.