Introduktion
Jag har haft turen att delta i två stora fysiologiska forskningsexpeditioner till extrema höjder. Den första var Silver Hut-expeditionen 1960-1961 under vilken en grupp fysiologer tillbringade flera månader på den mycket höga höjden 5 800 m (19 000 fot), och vi mätte de fysiologiska förändringar som inträffade under denna långa period. Ytterligare studier gjordes upp till 7 830 m. (25 700 fot). Det övergripande syftet var att fastställa de mekanismer genom vilka människor som normalt lever nära havsnivån reagerar på svår hypoxi under en längre period.
Den andra expeditionen var den amerikanska medicinska forskningsexpeditionen till Everest som ägde rum 1981. Här var det fysiologiska målet mycket annorlunda. Målet var att få fram de första människofysiologiska uppgifterna på toppen av Mount Everest (8 8 848 m, 29 028 fot) för att klargöra hur människor som normalt lever på havsnivå kan överleva den extrema hypoxi som råder på världens högsta punkt.
Silver Hut Expedition
Detta var Sir Edmund Hillarys hjärna som tillsammans med Tenzing Norgay var den förste personen som nådde toppen av Mount Everest sju år tidigare. Hillary hade samarbetat med Griffith Pugh, en höghöjdsfysiolog, vid denna första bestigning av Everest, och båda männen var intensivt intresserade av den acklimatiseringsprocess som gör det möjligt för människor från nära havsnivå att klättra till mycket höga höjder.
Mitt särskilda intresse gällde lungornas diffusionskapacitet. Barcroft (1925) hade föreslagit att träning på hög höjd skulle leda till arteriell hypoxemi på grund av diffusionsbegränsning över blodgasbarriären. För att testa detta ordnade vi så att expeditionsmedlemmarna arbetade med maximal kapacitet på en cykelergometer, och vi mätte den arteriella syremättnaden med hjälp av en nytillgänglig öronoximeter (West et al., 1962). Vi fann att det faktiskt skedde en markant minskning av den arteriella syremättnaden trots ökningen av den alveolära PO2 när arbetsnivån höjdes. Detta var ett starkt bevis för diffusionsbegränsning under dessa förhållanden med allvarlig hypoxi. Dubbla mätningar på mig själv visade en syremättnad på endast 33 % vid maximal ansträngning, vilket återspeglade mycket allvarlig hypoxemi.
Vi mätte också diffusionskapaciteten för kolmonoxid under expeditionens gång och visade att den knappast förändrades (West, 1962). Den lilla ökningen kunde förklaras av den polycytemi som utvecklades. Slutsatsen var att blodgasbarriärens egenskaper inte förändrades av långvarig exponering för svår hypoxi. Detta var den första tydliga demonstrationen av diffusionsbegränsning av syreöverföringen från lungan under svår träning på mycket hög höjd.
Senare under expeditionen gjordes mätningar av det maximala syreupptaget under träning på den extremt höga höjden 7 440 m (24 400 fot) (Pugh et al., 1964). Extrapolering av dessa data till Everest-toppens höjd tydde på att det skulle vara omöjligt att nå toppen utan extra syrgas. Blodundersökningar visade på en tydlig polycytemi hos de personer som levde på 5 800 meters höjd. Medelvärdet för hemoglobinkoncentrationen och hematokriten var 19,6 g/dl respektive 55,8 %. Det fanns belägg för att den initiala ökningen av hematokriten huvudsakligen orsakades av förlust av plasmavolym, men senare skedde en stor ökning av massan av röda blodkroppar. Elektrokardiogrammet hos personer som levde på 5 800 m höjd visade på en markant högerkammarhypertrofi, och i vissa spårningar fanns det en inversion av T-vågorna i bröstledningarna, vilket förmodligen tyder på allvarlig myokardiell hypoxi (Milledge, 1963). Mätningar av neuropsykometriska funktioner gjordes med hjälp av kortsortering, och man fann att sorteringshastigheten minskade men att försökspersonerna med ökad koncentration kunde sortera utan fel. Det förekom en allvarlig, obeveklig viktnedgång hos alla expeditionsmedlemmar när de levde på 5 800 meters höjd, med en hastighet på mellan 0,5 och 1,5 kg/vecka. Det allmänna intrycket var att det inte skulle vara möjligt för människor att leva på obestämd tid på denna mycket höga höjd (Pugh, 1962).
American Medical Research Expedition to Everest
Som nämnts ovan var syftet med denna expedition att klargöra hur människor kan tolerera hypoxi på världens högsta höjd. Anmärkningsvärt nog nådde två europeiska klättrare några månader innan expeditionen ägde rum för första gången Mount Everests topp utan att använda extra syrgas. Denna bedrift förvånade många fysiologer och väckte många frågor om hur det kunde göras.
Forskningsprogrammet var mycket omfattande och endast en kort sammanfattning kan ges här. Mätningar gjordes i baslägret på 5 400 m höjd, i det avancerade baslägret på 6 300 m höjd och i det högsta lägret på 8 050 m höjd. Vi hoppades också kunna göra några mätningar på Everests topp även om detta var mycket ambitiöst. Faktum är att när vi såg tillbaka på de sex expeditioner som föregått vår egen expedition hade inte en enda av dem nått toppen. Om vädret är dåligt kan man glömma det, och en kritisk faktor är om tillräckligt många expeditionsmedlemmar förblir tillräckligt friska trots den extrema höjden.
I baslägret mätte vi den ventilationsmässiga responsen på hypoxi, det vill säga i vilken utsträckning andningen ökar när försökspersonen utsätts för en lågt inspirerad syreblandning. Resultaten var slående. Det visade sig att den klättrare som nådde toppen först hade den högsta responsen, den som nådde toppen på andra plats hade den näst högsta responsen och den klättrare som nådde toppen på tredje plats hade den tredje högsta responsen (Schoene et al., 1984). Detta måste delvis vara en slump, men det tydde verkligen på att det fanns ett samband mellan i vilken utsträckning klättrare ökar sin ventilation och deras tolerans för extrema höjder. Orsaken till detta kommer att bli tydligare nedan.
Ett stort antal studier genomfördes i det avancerade baslägret men endast en, en neuropsykometrisk studie, kommer att sammanfattas här. Det är välkänt att hjärnan och det centrala nervsystemet är mycket känsliga för hypoxi. Om någon t.ex. faller i en simbassäng och räddas 5 eller 10 minuter senare kan han återupplivas framgångsrikt, men det centrala nervsystemet återhämtar sig aldrig helt och hållet. Det var därför inte förvånande att vi kunde visa förändringar i mätningar som korttidsminne och manipuleringsförmåga (som bestämdes med hjälp av ett test för att knacka med fingrarna) på mycket höga höjder. Detta var inte oväntat. När expeditionen återvände till nära havsnivå visade det sig dock att två av mätningarna förblev onormala. Dessa var korttidsminnet och fingertestet (Townes et al., 1984). Det stod därför klart att alla som återvänder från dessa extrema höjder sannolikt kommer att ha en viss kvarstående nedsättning av det centrala nervsystemet. Vi var en av de första grupperna som visade detta, men det har bekräftats många gånger sedan dess.
Några av de mest intressanta resultaten var de från toppmötet. Vi hade konstruerat och byggt en speciell anordning som gjorde det möjligt för klättraren att samla in den sista utandningsgasen efter en maximal utandning, det vill säga ett alveolärt gasprov. Över 34 prover, inklusive fyra från toppmötet, togs tillbaka till UC San Diego i gastäta burkar. När den alveolära PCO2-halten plottades mot det barometriska trycket, som sjönk med stigande höjd, fann man att PCO2-halten på toppmötet var 7-8 mm Hg. Detta var ett nästan otroligt lågt värde eftersom värdet på havsnivå är ca 40 mm Hg. Detta extremt låga värde understryker den enorma ökning av den alveolära ventilationen som är nödvändig på dessa extrema höjder (West et al., 1983).
När både den alveolära PO2 och PCO2 plottades mot det barometriska trycket framträdde en intressant bild (figur 1). Både PO2 och PCO2 sjönk när höjden ökade. Fallet i PO2 inträffade på grund av minskningen av luften runt klättraren till följd av minskningen av det barometriska trycket. Nedgången i PCO2 orsakades endast av klättrarens hyperventilation. Det visade sig att när höjden översteg ca 7000 m, skedde ingen ytterligare minskning av den alveolära PO2-nivån. Figuren visar att detta försvaras vid en nivå på cirka 35 mm Hg. Med andra ord, för att överleva på dessa enorma höjder måste man sätta upp en alveolär ventilation som driver ner PCO2 till cirka 8 mm Hg och därmed bevarar den alveolära PO2 på den mycket låga men livskraftiga nivån på cirka 35 mm Hg. Detta förklarar varför det i de tidigare nämnda mätningarna av den respiratoriska responsen på hypoxi fanns en korrelation mellan storleken på responsen och klättrarens tolerans för extrema höjder. Om man inte kan åstadkomma en hyperventilation som är tillräcklig för att driva ner den alveolära PCO2-halten till cirka 7-8 mm Hg kan man inte upprätthålla en livskraftig PO2-nivå i den alveolära gasen. Extrem hyperventilation är således ett av de viktigaste inslagen i det fysiologiska svaret på extremt hög höjd.
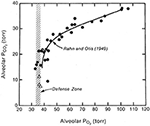
Figur 1. Värden för PO2 och PCO2 i alveolärgasen hos klättrare när de stiger från havsnivå (överst till höger) till Everest-toppen (nederst till vänster). PO2 sjunker på grund av det minskade barometriska trycket. PCO2 sjunker på grund av den ökande alveolära ventilationen. Ovanför en höjd som motsvarar en PCO2 på ca 20 mm Hg (ca 7000 m) sjunker PO2 inte längre. Med andra ord försvaras detta vid cirka 35 mm Hg. Detta kan endast ske om PCO2 kontinuerligt sänks ytterligare genom extrem hyperventilation när höjden stiger. Modifierat från Rahn och Otis (1949) och West et al. (1983).
Den extremt låga alveolära PCO2 föranleder frågan om vad som hände med det arteriella pH-värdet. Det är rimligt att anta att det arteriella och alveolära PCO2 är detsamma. Lyckligtvis tog två av klättrarna venöst blod från varandra på morgonen efter deras framgångsrika klättring till toppen, och basöverskottsvärdena kunde därför mätas. När dessa värden fördes in i Henderson-Hasselbalch-ekvationen var det resulterande arteriella pH-värdet mellan 7,7 och 7,8. Detta är en extrem grad av respiratorisk alkalos.
En intressant fråga är varför njurarna inte eliminerade bikarbonat för att utveckla en metabolisk kompensation för denna extrema alkalos och på så sätt föra pH närmare det normala. Detta är den vanliga reaktionen om en respiratorisk alkalos genereras, till exempel genom hyperventilation som ibland inträffar under hysteri. Orsaken till avsaknaden av metabolisk kompensation är inte helt klarlagd, men en möjlighet är att dessa bergsklättrare troligen var allvarligt utarmade på volym. Detta är ett vanligt inslag när man går på hög höjd och till exempel visade de klättrare som stannade i det avancerade baslägret på 6300 m höjd tecken på kronisk volymminskning. En ansvarig faktor på extrem höjd är förmodligen den enorma hyperventilationen, men även törsten är nedsatt.
De fysiologiska konsekvenserna av den allvarliga alkalosen är intressanta. Andra studier har visat att en ökad syreaffinitet hos hemoglobin ökar överlevnaden i en hypoxisk miljö. För många år sedan visades att däggdjur som vicuña och lama i de sydamerikanska Anderna har en ökad syreaffinitet (det vill säga de hade en vänsterförskjuten syrgasdissociationskurva) jämfört med däggdjur som lever på havsnivå (Hall, 1937). Klättrare på mycket hög höjd har alltså samma reaktion.
Det är också sant att om man tittar generellt i hela djurriket på organismer som lever i en hypoxisk miljö, har många av dem utvecklat en ökad syreaffinitet hos hemoglobinet. Ett av de bästa exemplen är det mänskliga fostret som på grund av en skillnad i hemoglobinets aminosyrasekvens har en markant ökad syreaffinitet med P50 på cirka 19 mm Hg jämfört med cirka 27 för en vuxen. Det mänskliga fostret har allvarlig hypoxemi enligt vuxenstandard med ett PO2 i den nedåtgående aorta på cirka 22 mm Hg, vilket till och med är lägre än hos en klättrare på Everest-toppen. Det är fascinerande att den framgångsrika klättraren har fördelen av en ökad syreaffinitet hos hemoglobinet. Detta underlättar laddningen av syre i lungkapillärerna. Man skulle kunna hävda att det också stör avlastningen av syre i kroppens periferi, men studier har visat att fördelen med belastning i lungan är större än nackdelen med avlastning i de perifera vävnaderna.
En intressant fråga är vad som är den maximala syreförbrukningen hos en klättrare på toppmötet. Som tidigare nämnts tyder tidigare mätningar som gjordes på extrem höjd under Silver Hut-expeditionen på att allt syre som finns tillgängligt på toppen skulle behövas för den basala syreuppdateringen, det vill säga för att hålla hjärtat pumpande och hjärnan aktiv. Det var omöjligt att placera en cykelergometer på toppen. Vi tog dock klättrare som var mycket väl acklimatiserade och lät dem trampa maximalt på en höjd av 6300 m samtidigt som de andades 14 % syre. Detta gav dem samma inspirerade PO2 som på toppen. Syreupptagningsnivån under dessa förhållanden var cirka 1 L/min, vilket är en uselt låg maximal syreförbrukning och motsvarar någon som går långsamt på nivån. Det är dock tydligen precis tillräckligt för att en klättrare ska kunna nå toppen.
Författarbidrag
JW utarbetade manuskriptet, läste och godkände den inlämnade versionen.
Intressekonfliktförklaring
Författaren förklarar att forskningen utfördes i avsaknad av kommersiella eller ekonomiska relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Barcroft, J. (1925). Blodets andningsfunktion. Del 1 Lärdomar från höga höjder. Cambridge: Cambridge University Press.
Google Scholar
Hall, F. G. (1937). Djurens anpassning till hög höjd. J. Mammol. 18, 469-472. doi: 10.2307/1374337
CrossRef Full Text | Google Scholar
Milledge, J. S. (1963). Elektrokardiografiska förändringar på hög höjd. Br. Heart J. 15, 291-298. doi: 10.1136/hrt.25.3.291
CrossRef Full Text | Google Scholar
Pugh, L. G. (1962). Fysiologiska och medicinska aspekter av Himalaya- och vetenskapsexpeditionen 1960-61. Br. Med. J. 2, 621-633. doi: 10.1136/bmj.2.5305.621
CrossRef Full Text | Google Scholar
Pugh, L. G., Gill, M. B., Lahiri, S., Milledge, J. S., Ward, M. P. och West, J. B. (1964). Muskelträning på stora höjder. J. Appl. Physiol 19, 431-440. doi: 10.1152/jappl.1964.19.3.431
CrossRef Full Text | Google Scholar
Rahn, H., and Otis, A. B. (1949). Människans andningsrespons under och efter acklimatisering till hög höjd. Am. J. Physiol. 157, 445-462. doi: 10.1152/ajplegacy.1949.157.3.445
PubMed Abstract | CrossRef Full Text | Google Scholar
Schoene, R. B., Lahiri, S., Hackett, P. H., Peters, R. M., Milledge, J. S., Pizzo, C. J., et al. (1984). Förhållandet mellan den hypoxiska ventilationsresponsen och träningsprestationen på Mount Everest. J. Appl. Physiol. 56, 1478-1483. doi: 10.1152/jappl.1984.56.6.1478
CrossRef Full Text | Google Scholar
Townes, B. D., Hornbein, T. H., and Grant, I. (1984). ”Human cerebral function at extreme altitude,” in High Altitude and Man, eds J. B. West and S. Lahiri (Bethesda, MD: American Physiological Society), 31-36.
Google Scholar
West, J. B. (1962). Lungans diffusionskapacitet för kolmonoxid på hög höjd. J. Appl. Physiol. 17, 421-426. doi: 10.1152/jappl.1962.17.3.421
CrossRef Full Text | Google Scholar
West, J. B., Hackett, P. H., Maret, K. H., Milledge, J. S., Peters, R. M., Pizzo, C. J., et al. (1983). Lungegasutbyte på toppen av Mount Everest. J. Appl. Physiol. 55:678-687. doi: 10.1152/jappl.1983.55.3.678
CrossRef Full Text | Google Scholar
West, J. B., Lahiri, S., Gill, M. B., Milledge, J. S., Pugh, L. G. C. E. och Ward, M. P. (1962). Arteriell syresättning under träning på hög höjd. J. Appl. Physiol. 17, 617-621. doi: 10.1152/jappl.1962.17.4.617
CrossRef Full Text | Google Scholar