- Verbindung C1 kann DNA spalten, die eine AP-Stelle enthält
- Einige strukturelle Einheiten in C1 sind für die Katalyse wichtig
- DNA-Struktur moduliert die C1-katalysierte Spaltung
- Die Abspaltung von DNA an AP-Stellen durch C1 erfolgt über ein Zwischenprodukt, an dem das sekundäre Amin beteiligt ist
- C1 erhöht die thermische Stabilität von DNA, die ein Analogon einer AP-Stelle enthält
- C1 hat eine höhere Affinität für DNA, die eine AP-Stelle enthält
- Schlussfolgerungen
Verbindung C1 kann DNA spalten, die eine AP-Stelle enthält
Die erste Beobachtung, die das Vorhandensein einer ungewöhnlichen Aktivität in einer Untergruppe der untersuchten kleinen Moleküle nahelegte, wurde gemacht, als C1 (Abb. 1a) als potenzieller Inhibitor von hOGG1 getestet wurde. Insbesondere führte die Zugabe dieser Verbindung zu Reaktionen von hOGG1 mit einem fluoreszenzmarkierten Duplex-Oligodeoxynukleotid, das ein ortsspezifisches 8-oxo-dG-Addukt enthält (Abb. 1b), zu einem zusätzlichen Produkt, das schneller wanderte als das β-Eliminierungsprodukt; hOGG1 allein ergab nur das β-Eliminierungsprodukt (Abb. 1c). Bei der Inkubation von 8-oxo-dG-haltiger DNA mit C1 allein wurden keine Nicked-Produkte beobachtet.

AP-Stelle-haltige DNA wurde durch Behandlung der entsprechenden dU-haltigen DNA mit UDG erhalten. (a) Struktur eines repräsentativen Katalysators für die DNA-Spaltung. (b) DNA-Substrate. (c) Spaltung von 8-oxo-dG-haltiger DNA (250 nM) in Gegenwart von hOGG1 (100 nM) und C1 (10 μM). (d) Spaltung von AP-stellenhaltiger DNA (250 nM) in Gegenwart von hOGG1 (50 nM) und C1 (10 μM). (e) Spaltung von AP-stellenhaltiger DNA (250 nM) in Gegenwart von hNEIL1 (50 nM) und C1 (10 μM). (f) Spaltung von AP-Stellen-haltiger DNA (2 μM) durch C1. Der Prozentsatz der Produkte wurde um die spontane Spaltung korrigiert. Die Reaktionen wurden bei 37 °C für 30 min (b-d) oder 16 h (e) durchgeführt.
Es wurden mehrere mechanistische Möglichkeiten angenommen, die die Bildung des neuen Produkts erklären könnten. 1) In Gegenwart von C1 hatte hOGG1 die Fähigkeit erworben, zusätzlich zu seinen Glykosylase- und β-Eliminierungs-AP-Lyase-Funktionen eine δ-Eliminierungsreaktion zu katalysieren; 2) C1 könnte das β-Eliminierungsprodukt in das δ-Eliminierungsprodukt umwandeln; und 3) AP-Stellen, die in der Glykosylasereaktion gebildet werden, könnten als Substrat für eine C1-katalysierte β,δ-Eliminierungsreaktion dienen. Um diese Möglichkeiten zu untersuchen, wurde Duplex-DNA, die eine einzelne ortsspezifische AP-Stelle enthält, durch Behandlung der entsprechenden dU-haltigen DNA mit Uracil-DNA-Glykosylase (UDG) erzeugt (Abb. 1b) und mit C1 oder hOGG1, entweder einzeln oder in Kombination, zur Reaktion gebracht. Wie bei dem 8-oxo-dG-haltigen Oligodeoxynukleotid erzeugte hOGG1 das erwartete β-Eliminierungsprodukt, während die Zugabe von C1 ein Gemisch aus zwei Produkten ergab (Abb. 1d). Dieses Produktgemisch wurde auch gebildet, wenn das AP-haltige Oligodeoxynukleotid mit C1 allein inkubiert wurde. Während die Position der langsamer wandernden Produktbande dem β-Eliminierungsprodukt von hOGG1 entsprach (Abb. 1d), wanderte die schneller wandernde Produktbande mit dem bekannten δ-Eliminierungsprodukt von hNEIL1 mit (Abb. 1e). Somit fördert C1 die β- und δ-Eliminierungsreaktionen an AP-Stellen.
Die Spaltungsreaktion durch C1 an AP-Stellen enthaltender DNA war konzentrationsabhängig (Abb. 1e und ergänzende Abbildung S1), wobei niedrige mikromolare Konzentrationen ausreichten, um die Produkte zu beobachten. Ähnlich wie die zuvor entwickelten „künstlichen Nukleasen „26 zeigte C1 eine Turnover-Katalyse auf dem DNA-Substrat: 1 pmol C1 erzeugte ~1,4 pmol Produkte in 16 Stunden (Abb. 1f). Die Schnittgeschwindigkeit von C1 an dieser DNA lag also bei ~1,5 × 10-3 min-1 oder höher. Der Vergleich von C1 mit den derzeit erhältlichen AP-Lyase-Reagenzien Spermin17 und KWKK-Peptid23 zeigte, dass der neu identifizierte Katalysator mindestens 100-mal effizienter ist (ergänzende Abbildung S1).
Einige strukturelle Einheiten in C1 sind für die Katalyse wichtig
Analoga von C1 wurden untersucht, um den Beitrag spezifischer struktureller Merkmale zur Strangspaltungschemie zu bestimmen. Zunächst untersuchten wir die Fähigkeit einfacher, handelsüblicher Amine ohne den Indolinon-Pyrrol-Anteil C2 (Abb. 2) und CS1-CS3 (ergänzende Abbildung S2), die DNA an einer AP-Stelle einzuschneiden. Die Bildung von Schnittprodukten, wenn überhaupt, lag bei 10 μM unterhalb der Nachweisgrenze. Der Indolinon-Pyrrol-Anteil ist also wichtig für die Reaktion. Es ist möglich, dass die Indolinon-Pyrrol-Untereinheit an die AP-Stelle29 oder die Nebenfurche bindet und das sekundäre Amin positioniert, um die Strangspaltung über ein kovalentes Zwischenprodukt mit der AP-Stelle zu katalysieren. Die Verbindungen C3 (Abb. 2) und CS4 (ergänzende Abbildung S2), die die Indolinon-Pyrrol-Einheit enthielten, denen aber das sekundäre Amin fehlte, waren ebenfalls völlig inaktiv. Diese Daten deuten stark auf die Rolle des Amins als reaktive funktionelle Gruppe hin. Der Ersatz der Methoxygruppe am Pyrrol durch andere Substitutionen, z. B. in C4 und C5 (Abb. 2), führte zu einer geringeren Aktivität. Die Reaktion wurde auch durch einen Substituenten an der Aminogruppe moduliert, so dass eine Erhöhung der sterischen Anforderungen des sekundären Amins die Menge des beobachteten Produkts verringerte (vergleiche C1, C6 und C7 (Abb. 2) und C4, CS5 und CS6 (ergänzende Abbildung S2)). Diese Struktur-Aktivitäts-Analysen zeigten, dass mehrere der einzelnen Komponenten für die Spaltung notwendig, aber nicht ausreichend sind und dass diese Komponenten zusammenwirken müssen, um die Strangspaltung zu bewirken.

(a) Strukturen der repräsentativen Verbindungen. (b) DNA-Substrat. (c) Test auf die Fähigkeit der repräsentativen Verbindungen (10 μM), AP-haltige DNA (250 nM) einzuschneiden. Die Reaktionen wurden bei 37 °C für 30 Minuten durchgeführt.
DNA-Struktur moduliert die C1-katalysierte Spaltung
Die Bedeutung der Indolinon-Pyrrol-Einheit für die Strangspaltungs-Chemie hat nahegelegt, dass die Struktur des DNA-Substrats die C1-vermittelte Spaltung beeinflussen könnte. Um diese Zusammenhänge zu untersuchen, wurden die anfänglichen Reaktionsraten für eine einzelsträngige DNA (Sequenz wie in Abb. 1a) und die entsprechenden doppelsträngigen DNAs gemessen, die entweder A, C, G oder T gegenüber der AP-Stelle enthielten. Das homopolymere Oligodeoxynukleotid, 5′-TAMRA-(T)5-AP-(T)11-3′, wurde ebenfalls als eindeutige einzelsträngige DNA-Struktur untersucht. Diese Daten zeigten, dass die Strangspaltung zwar auch im Zusammenhang mit einzelsträngiger DNA stattfinden kann, dass aber doppelsträngige DNAs die bevorzugten Substrate für C1 sind (Abb. 3). Es wurde auch festgestellt, dass die Art der Base, die der AP-Stelle gegenüberliegt, die Katalyse moduliert, wobei die Geschwindigkeit der Hydrolyse der AP-Stelle gegenüber Pyrimidinen schneller ist als gegenüber Purinen. Diese Beobachtungen stimmen mit dem Vorschlag überein, dass die Indolinon-Pyrrol-Untereinheit mit der DNA interagiert, um einen leeren Raum an der AP-Stelle zu besetzen. Bemerkenswert ist, dass die für die doppelsträngige DNA mit einem C gegenüber der AP-Stelle gemessene Anfangsrate sehr nahe an der für dieses Substrat unter Bedingungen mit begrenzter C1-Konzentration beobachteten Rate lag (Abb. 1f).

Die Reaktionen wurden bei 37 °C mit 250 nM DNA und 5 μM C1 durchgeführt. Die mittleren Anfangsraten mit den jeweiligen Standardabweichungen wurden aus drei unabhängigen Experimenten mit der Software KaleidaGraph 4.1 (Synergy Software) berechnet. Die P-Werte wurden mit dem Students‘ t-Test berechnet.
Die Abspaltung von DNA an AP-Stellen durch C1 erfolgt über ein Zwischenprodukt, an dem das sekundäre Amin beteiligt ist
Um das kovalente Zwischenprodukt mit der ringgeöffneten, aldehydischen Form der Desoxyribose zu testen, wurde ein 32P-markiertes Oligodeoxynukleotid, das eine AP-Stelle enthält (Abb. 4a), mit C1 inkubiert. 4a) mit C1 in Gegenwart von NaB(CN)H3 inkubiert. Während dieses Reduktionsmittel langsam mit der AP-Läsion reagiert, fängt es das Imin- oder Iminium-Ionen-Konjugat effizient ein. In einer Kontrollreaktion mit dem AP-haltigen Oligodeoxynukleotid und NaB(CN)H3 (Abb. 4b, Spur 5) wies ein kleiner Teil (~3 %) der DNA eine verringerte Mobilität auf. Dieses Produkt mit geringer Häufigkeit wird häufig in den Trapping-Reaktionen23 beobachtet und stellt wahrscheinlich einen Komplex mit Tris-Molekülen dar, wie zuvor in Reaktionen mit dem Malondialdehyd-Pyrimidopurinon-DNA-Addukt30 gezeigt wurde. In einer positiven Kontrollreaktion mit dem Lysin-Tryptophan-Lysin-Lysin (KWKK)-Peptid23 wurde das Imin-Zwischenprodukt eingefangen, wie die Verschiebung der DNA-Mobilität zeigt (Abb. 4b, Lane 3). In Gegenwart von NaB(CN)H3 und C1 bildete der Großteil der DNA (~80 %) einen Komplex, der als Spezies mit verringerter Mobilität auftrat (Abb. 4b, Lane 4). Die Komplexbildung war nicht auf eine unspezifische Bindung von C1 an die DNA zurückzuführen, da keine Verschiebung beobachtet wurde, als das entsprechende dU-haltige Oligodeoxynukleotid unter identischen Bedingungen getestet wurde (Abb. 4b, Spur 1). Diese Daten stimmten mit der Hypothese überein, dass das Reagenz in der Lage war, ein kovalentes Iminium-Ionen-Zwischenprodukt zu bilden, das dann eine Seitengruppe für eine Protonenabstraktion vom Zuckerring positioniert.

(a) DNA-Substrate. (b) Cyanoborhydrid-Trapping eines Komplexes zwischen C1 und AP-haltiger DNA. (c) CID-Fragmentierung des reduzierten Iminium-Ionen-Zwischenprodukts. (d) Fragmentierung des reduzierten C1-Desoxyribose-Konjugats nach enzymatischem Verdau.
Das reduktive Einfangen von C1 wurde unter Verwendung von unmarkiertem AP-Stellen-haltigem Oligodeoxynukleotid (Abb. 4a) wiederholt, und das Produkt wurde mittels Massenspektrometrie (MS) analysiert. Die Analyse ergab eine Masse, die mit dem reduzierten kovalenten DNA-C1-Komplex übereinstimmt (m/z 1134,64 für -2). Die kollisionsinduzierte Dissoziation (CID) dieses Ions führte zu einem vollständigen Satz von a-B- (Base) und w-Ionen, die mit dem reduzierten Iminium-Ion zwischen C1 und der AP-Stelle übereinstimmen (ergänzende Abbildung S3). In der a-B-Ionenreihe geht die Fragmentierung der C3′-O-Bindung normalerweise mit dem neutralen Verlust der Nukleobase einher. Es wurde erwartet, dass die reduzierte Bindung zwischen C1 und der AP-Stelle weniger labil ist, weshalb wir sowohl das a4-Ion (m/z 1330,1) als auch das a4-B-Ion (m/z 1005,6) beobachteten. Das Oligodeoxynukleotid wurde auch enzymatisch verdaut und mittels MS analysiert (Abb. 4c und ergänzende Abbildung S4). Es wurde ein Verdauungsprodukt mit einer Masse beobachtet, die mit dem reduzierten C1-Desoxyribose-Konjugat übereinstimmt (m/z 444,19); die Fragmentierung dieses Produktions ergab ein Tochterion mit m/z 295,09, das aus dem neutralen Verlust von N-Methylamino-2-desoxyribitol resultierte (Abb. 4d und ergänzende Abbildung S4). Dieses Produkt war identisch mit dem Produkt aus der reduktiven Aminierungsreaktion von 2-Desoxyribose und C1. Diese Untersuchungen zeigen, dass C1 AP-Stellen enthaltende DNA durch kovalente Katalyse unter Beteiligung des sekundären Amins spaltet.
C1 erhöht die thermische Stabilität von DNA, die ein Analogon einer AP-Stelle enthält
Die Wirkung von C1 wurde auf die thermische Stabilität von DNA untersucht, die ein Tetrahydrofuran (THF) enthält, ein strukturelles Analogon einer AP-Stelle, das nicht in der Lage ist, die β-Eliminierungsreaktion zu durchlaufen31. Die Tm der Duplex-DNA, die THF gegenüber T (T:THF) enthält, stieg bei Zugabe von 1 Äquivalent C1 um 6,3 °C, während die Tm der Kontroll-DNA, die ein T:A-Paar enthält, weniger betroffen war und nur um 1 °C anstieg (Abb. 5). Während die Schmelzkurve von T:THF mit C1 auf eine thermische Stabilisierung durch C1 hindeutet, verbreitert die Anwesenheit von C1 auch die Schmelzkurve, was auf einen weniger kooperativen Schmelzübergang hinweist. Insgesamt deuten diese Beobachtungen darauf hin, dass C1 spezifisch an die DNA an der AP-Stelle bindet und die DNA-Helix lokalisiert stabilisiert. Dies steht im Einklang mit dem vorgeschlagenen Modell der Wechselwirkungen zwischen C1 und der DNA, bei dem der Indolinon-Pyrrol-Anteil den verfügbaren Platz an der AP-Stelle einnimmt.
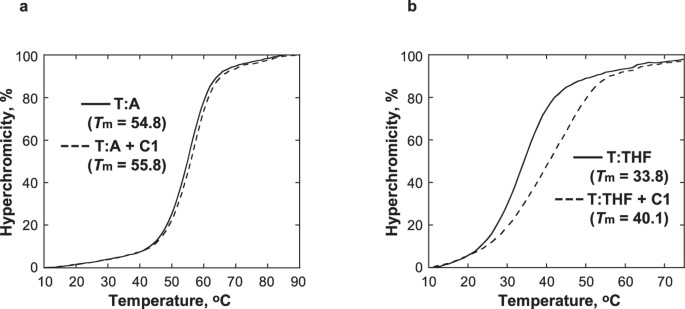
Die Schmelztemperaturkurven von T:A (a) und T:THF (b) wurden mit 5 μM DNA in 10 mM Natriumphosphatpuffer, pH 7.0, 100 mM NaCl und 1 mM EDTA in Abwesenheit (durchgezogene Linie) und Anwesenheit (gestrichelte Linie) von 5 μM C1.
C1 hat eine höhere Affinität für DNA, die eine AP-Stelle enthält
Um den Bindungsmodus und die Affinität von C1 zu AP-Stellen-haltiger DNA zu untersuchen, wurden zirkuläre Dichroismus (CD)-Analysen sowohl mit THF-haltigen als auch mit Kontroll-T:A-Duplex-Oligodeoxynukleotiden durchgeführt (Abb. 6). Die induzierte CD (ICD) von C1 bei Wechselwirkung mit dem T:THF-Duplex wurde als starkes Exzitonensignal beobachtet, das eine bisignierte Form mit positiven und negativen Banden relativ zum Absorptionsmaximum des freien C1 aufweist. Dies deutet im Allgemeinen auf die Bildung dimerer Komplexe oder von Komplexen höherer Ordnung hin, entweder im Modus der Groove-Bindung oder der externen Stacking-Bindung32. Die Titration von C1 mit steigenden T:THF-Konzentrationen (Abb. 6a und ergänzende Abbildung S5) zeigte sowohl unspezifische als auch spezifische Wechselwirkungen. Zu Beginn der Titration ist C1 im Überschuss vorhanden und unspezifische Wechselwirkungen werden bevorzugt. Nach Zugabe von T:THF stieg die ICD bei ~488 nm an. Bei einer DNA-Konzentration von etwa 2 μM verschob sich die ICD-Bande zu einer niedrigeren Energie (~495 nm) und nahm dann bis zum Äquivalenzpunkt ab, wo sie im DNA-Überschuss auf ein Plateau stieß, was auf eine spezifische Wechselwirkung hinweist. Die Titration von C1 mit der Kontroll-DNA T:A (Abb. 6a und ergänzende Abbildung S5) zeigte keine positive ICD-Bande um 495 nm, was auf das Fehlen spezifischer Wechselwirkungen hinweist. Die Vorwärtstitration von T:THF zeigte ebenfalls Hinweise auf unspezifische und spezifische Wechselwirkungen (Abb. 6b). Bei niedrigen C1-Konzentrationen mit einem Überschuss an DNA lag die ICD-Bande bei 495 nm (spezifisch); mit zunehmender C1-Konzentration im Verhältnis zur DNA nahm die ICD-Intensität zu und verschob sich zu einer höheren Energie (488 nm), was auf eine unspezifische Bindung hindeutet, wie sie bei hohen Ligandenkonzentrationen erwartet wird. Die beobachtete hypsochrome Verschiebung könnte auf eine veränderte oder besser definierte Konformation von C1 bei spezifischer Bindung an die AP-Stelle hinweisen. Die Titration der Kontroll-DNA ohne AP-Stelle zeigte ebenfalls eine ICD und Hinweise auf eine unspezifische Bindung (Abb. 6c). Es ist erwähnenswert, dass die Bande bei 450-520 nm nur bei einem 3-fachen Überschuss von C1 in Bezug auf die DNA auftrat, so dass dieser Wellenlängenbereich die beste Wahl für die Konstruktion der Bindungskurven war. Die Bindungsisothermen wurden anhand eines einfachen bimolekularen Bindungsmodells konstruiert und analysiert, wie bereits veröffentlicht32. Die nichtlinearen Gleichungen, die sich aus den 500-nm-Isothermen ergaben (Abb. 6d), ergaben eine Dissoziationskonstante (KD) von 64 μM für die T:A-DNA-Kontrolle (unspezifische Bindung) und 29 μM für die T:THF-DNA (spezifische Bindung); die KD von C1 mit THF-haltigem Duplex wurde nach Subtraktion zur Korrektur des unspezifischen Bindungsbeitrags und möglicher Ligand-Ligand-Wechselwirkungen auf 22 μM berechnet (Tabelle 1). Die bei 495 nm berechneten KD-Werte lagen nur geringfügig außerhalb der Fehler der bei 500 nm berechneten Werte (Tabelle 1).

(a) Titrationskurven der ICD-Intensität bei 500 nm unter Verwendung einer konstanten C1-Konzentration (20 μM) und steigender DNA-Konzentrationen (Rücktitration). (b) CD-Spektren für T:THF und (c) T:A unter Verwendung einer konstanten DNA-Konzentration (10 μM) und steigender Konzentrationen von C1 (Vorwärtstitration). (d) Titrationskurven der ICD-Intensität bei 500 nm gegen den Logarithmus der C1-Konzentration für T:THF und T:A (abgeleitet aus den Daten in den Feldern b und c). (e) Titrationskurven der ICD-Intensität bei 500 nm gegen den Logarithmus der C6-Konzentration für T:THF und T:A (abgeleitet aus den Daten in der ergänzenden Abbildung S6). (d) Titrationskurven der ICD-Intensität bei 500 nm gegen den Logarithmus der C6-Konzentration für T:AP und T:A (abgeleitet aus den Daten in der ergänzenden Abbildung S6).
Ferner wurde die Möglichkeit in Betracht gezogen, dass die Affinität von C1 für DNA, die eine natürliche AP-Stelle anstelle von THF enthält, anders sein könnte. Um dieser Frage nachzugehen, haben wir die strukturell verwandte, aber inaktive Verbindung C6 (Abb. 2) in CD-Analysen mit DNA getestet, die entweder THF oder eine AP-Stelle enthält, die durch UDG-Behandlung aus dU erzeugt wurde. Die CD-Spektren, die für C6 in Vorwärts-Titrationsexperimenten mit der T:THF- und T:A-DNA beobachtet wurden (ergänzende Abbildung S6), waren im Allgemeinen ähnlich wie die für C1 beobachteten (Abb. 6b,c). Die Bindungsisothermen wurden konstruiert (Abb. 6e) und wie oben beschrieben analysiert. Die KD von C6 mit THF-haltigem Duplex, korrigiert um die unspezifischen Wechselwirkungen, wurde auf 28 μM berechnet (Tabelle 1), was nur ~25 % höher ist als die für C1 berechnete KD. Somit scheint C6 ein geeignetes Modell für die Untersuchung der Wechselwirkungen von C1 mit DNA zu sein. Die Vorwärtstitration von C6 wurde dann mit DNA durchgeführt, die die von UDG abgeleitete AP-Stelle enthielt (ergänzende Abbildung S6). Die Analyse der Bindungsisothermen ergab, dass die Affinität von C6 zu dieser DNA im Wesentlichen identisch mit der THF-haltigen DNA war (Abb. 6f und Tabelle 1). In Anbetracht der Tatsache, dass Desoxyribose an einer natürlichen AP-Stelle überwiegend in der ringgeschlossenen, THF-ähnlichen Form vorliegt31 , war das letztgenannte Ergebnis nicht überraschend. Es kann davon ausgegangen werden, dass die Affinität von C1 zu AP-haltiger DNA vor der Bildung eines kovalenten Zwischenprodukts vergleichbar mit der für THF-haltige DNA gemessenen Affinität ist. Die stärkere Bindung von C1 an DNA, die eine AP-Stelle enthält, im Vergleich zur Kontroll-DNA stimmt also mit den Daten zur thermischen Stabilität überein und unterstützt den Vorschlag, dass die Indolinon-Pyrrol-Untereinheit einen leeren Raum an der AP-Stelle der DNA besetzt. Es ist wahrscheinlich, dass die Affinität von C1 für die AP-Stelle zu seiner Lyaseaktivität beiträgt und einen klaren Vorteil gegenüber C2, CS1, CS2 und CS3 bietet, denen die Indolinon-Pyrrol-Einheit fehlt (Abb. 2 und ergänzende Abbildung S2).
Schlussfolgerungen
Die in dieser Studie beschriebenen AP-DNA-Strangspaltungskatalysatoren stellen eine neue Klasse von Verbindungen dar, die zur Spaltung von AP-Stellen unter physiologischen Bedingungen eingesetzt werden können. Auf der Grundlage dieser Kernstruktur können Moleküle entwickelt werden, um die Selektivität der AP-Stellen-Spaltung gegenüber verschiedenen Basen und möglicherweise für verschiedene Sequenzkontexte zu verbessern. Aufgrund der jüngsten Fortschritte bei der Entwicklung von Hochdurchsatz-Screening-Techniken könnten diese Verbindungen rasch zu effizienten Reagenzien für die Spaltung von AP-Stellen weiterentwickelt werden, die durch Depurinierung oder den BER-Weg in Zellen und Organismen entstehen. Die optimierten Versionen von C1 könnten auch therapeutisch eingesetzt werden, insbesondere in Kombination mit vielen gängigen Krebsmedikamenten, die entweder die DNA schädigen (z. B. Alkylierungsmittel33) oder auf die DNA-Reparatur abzielen (z. B. PARP34,35 oder AP-Endonuklease36-Inhibitoren). Es ist erwähnenswert, dass die 3′-Enden, die durch β- oder β,δ-Eliminierung entstehen, von DNA-Polymerasen ohne vorherige Reparatur37 nicht genutzt werden können. Die biologischen Folgen der Umwandlung von AP-Stellen in solche DNA-Strangbrüche dürften komplex sein und je nach der zellulären Fähigkeit, verschiedene Arten von DNA-Schäden zu reparieren oder zu tolerieren, variieren. Zu den möglichen Folgen gehören eine erhöhte therapeutische Wirksamkeit von Krebsbehandlungen (effizientere Zellabtötung) und eine geringere medikamenteninduzierte Mutagenese. Die Spaltung der DNA an AP-Stellen könnte sich als besonders vorteilhaft für die Behandlung von Krebsarten erweisen, die Defekte in den Mechanismen zur Reparatur von DNA-Strangbrüchen aufweisen, wie z. B. BRCA-defiziente Krebsarten34,35.